
ABSTRACT
Local anesthetics inhibit cell proliferation and induce apoptosis in various cell types. Ropivacaine, a unique, novel tertiary amine-type anesthetic, was shown to inhibit the proliferation of several cell types including keratinocytes. We found that Ropivacaine could inhibit the proliferation and induce apoptosis in an immortalized human keratinocyte line, HaCaT, in a dose- and time-dependent manner and with the deprivation of serum. The dose-dependent induction of apoptosis by ropivacaine was demonstrated by DNA fragmentation analysis and the proteolytic cleavage of a caspase-3 substrate — poly (ADP-ribose) polymerase (PARP). In addition, ropivacaine downregulated the expression of clusterin/apoliporotein J, a protein with anti-apoptotic properties, in a dose-dependent manner, which well correlated with the induction of apoptosis of HaCaT cells. To investigate the role of clusterin/apoliporotein J in ropivacaine-induced apoptosis, HaCaT cells overexpressing clusterin/apoliporotein J were generated and compared to cells expressing the well established anti-apoptotic Bcl-2 protein. Ectopic overexpression of the secreted form of clusterin/apoliporotein J or Bcl-2 decreased the sensitivity of HaCaT cells to toxic effects of ropivacaine as demonstrated by DNA fragmentation, the proteolytic cleavage of PARP and by a reduction in procaspase-3 expression. Furthermore, the downregulation of endogenous clusterin/apolipoprotein J levels by ropivacaine suggested that this might be one mechanism by which ropivacaine induced cell death in HaCaT cells. In conclusion, the ability of ropivacaine to induce antiproliferative responses and to suppress the expression of the anti-apoptotic protein clusterin/apolipoprotein J, combined with previously reported anti-inflammatory activity and analgesic property of the drug, suggests that ropivacaine may have potential utility in the local treatment of tumors.
Similar content being viewed by others

INTRODUCTION
Local anesthetics were shown to affect various neuronal as well as non-neuronal cellular activities and to induce cytotoxic effects and apoptosis. Ropivacaine (Ropivacaine hydrochloride; NaropeineTM), a unique, novel amide-type local anesthetic, is recently introduced to the market available for clinical use. Ropivacaine has been developed to replace bupivacaine as a reversible blocker of nerve fibers with higher efficacy/toxicity ratio 1, 2. In addition to its action as an effective analgesic/ anesthetic, ropivacaine possesses anti-inflammatory properties. It was demonstrated to inhibit eicosanoid release (5-lipoxygenase products) in human granulocytes and endothelial cells in vitro 3, block inflammatory leukocyte adhesion, CD11b/CD18 up-regulation and L-selectin shedding induced by tumor necrosis factor alpha (TNF-α) 4, 5 and decrease the vascular permeability in vivo, as well as the release of inflammatory mediators such as leukotriene B4 and 5-HETE 5. Ropivacaine was also demonstrated to be involved in inhibiting human platelet aggregation, microsomal lipid peroxidation and the expression of cytokine and adhesion molecules induced by lymphocyte activation 6. Furthermore, ropivacaine also affected mitochondrial function and energy metabolism in Ehrlich ascites tumor cells 7 and in rat liver mitochondria 8.
There are relatively fewer in vitro studies investigating the effects of local anesthetics on cell proliferation, apoptosis and gene expression 9, 10, 11, 12, 13, 14. It was reported that ropivacaine was shown to inhibit the proliferation of non-transformed cultured adult human fibroblasts, endothelial cells, keratinocytes 12 and primary rat fibroblast 6, as well as that of the transformed monkey kidney Vero cells 6 and human colon cell lines 15. Despite these studies, however, little is known about the mechanisms and signaling pathways through which ropivacaine inhibits cell proliferation or if, under certain conditions, induces apoptosis and alters gene expression.
Bcl-2 and clusterin/apolipoprotein J (ApoJ) are both well known for their functions in inhibiting apoptosis and increasing cell survival. Although the anti-apoptotic role of Bcl-2 to various toxic agents and stress stimuli is well-established 16, that of clusterin/ApoJ is still under investigation 17, 18. Clusterin is ubiquitously expressed, implicated in a number of physiological functions and is induced in diverse diseases and stress stimuli such as heat shock and oxidative stress, cytokines and growth factors, chemotherapeutic agents, ultraviolet and ionizing radiation. This led to suggestion that it may play a role in membrane lipid cycling, apoptotic cell death, and as a stress-induced secreted chaperone protein 17, 18. Overexpression of the secreted form of clusterin/ApoJ in human tumor cells would induce resistance to TNFα- 19, 20, Fas- 21, oxidative stress- 22 or chemotherapy-mediated apoptosis 23. Antisense technology used to inhibit endogenous clusterin in A431 cells confirmed a phenotype sensitive to heat shock and oxidative stress leading to their apoptosis 24. Similarly, antisense oligonucleotides targeting the clusterin gene in various tumor cell lines resulted in the inhibition of tumor growth and conferred chemosensitivity 25, 26, 27.
In this study we show that ropivacaine inhibited the proliferation and induced apoptosis of a human keratinocyte cell line (HaCaT) in a dose- and time-dependent manner, an observation correlated with the downregulation of clusterin/ApoJ. Overexpression of human clusterin/ApoJ or Bcl-2 protected HaCaT cells from ropivacaine-induced apoptosis.
MATERIALS AND METHODS
Cell culture
A spontaneously immortalized human skin keratinocyte line (HaCaT) which was obtained from Drs. N. Fusenig and P. Boukamp (German Cancer Research Center, German Cancer Research Center, Heidelberg, Germany) was cultured in DMEM supplemented with 10% (v/v) fetal calf serum (FCS), 2 mM L-glutamine, 100 units/ml penicillin and 100 μg/ml streptomycin (Biochrom, Germany) in a humidified incubator at 37°C.
Expression vectors and DNA transfections
By using the DMSO-polybrene method, HaCaT cells were transfected with pcDNA3.1B (Invitrogen, Life Technologies, USA), pcDNA3.1/hApoJ that carries the human clusterin/ApoJ cDNA and neo 28 or pcDNA3/hBcl-2 that carries human Bcl-2 cDNA and neo 29, respectively. After being selected in 500 μg/ml G418 for 3 weeks, several stably transfected HaCaTBcl-2 or HaCaTApoJ clones were picked out and subjected to immunoblotting for the expression of clusterin/ApoJ. HaCaTBcl-2 clone 2 (HaCaTBcl-2/2) and HaCaTApoJ clone 2 (HaCaTApoJ/2) exhibited high expression and were selected for further analysis. HacaTneo cells were used as G418-resistant cell population.
Ropivacaine treatment of HaCaT cells and analysis of cell proliferation
Untransfected HaCaT cells, HaCaTNeo, HaCaTBcl-2/2 and HaCaTApoJ/2 cells were plated in 24-well plates in duplicate at a density of 1×105 cells per well and allowed to grow for 36 h in complete DMEM. Then the medium was aspirated and the cells were treated with 0, 0.25, 0.5, 1.0, 2.0, 3.0 and 5.0 mM ropivacaine (ropivacaine hydrochloride; NaropeineTM) (Astra Zeneca, SA, Sweden) for 24 or 48 h in FCS free DMEM. The cells were harvested at the indicated times and counted on a Neubauer chamber. Cell viability was determined by the trypan blue exclusion assay. To study cell morphology changes, untreated and ropivacaine-treated cells were observed under a Zeiss Model Axiovert S100 Zeiss Model Axiovert S100, Germany and photographed.
DNA fragmentation assay
Untransfected HaCaT, HaCaTNeo, HaCaTBcl-2/2 and HaCaTApoJ/2 cells seeded at a density of 1-2×106 cells per 10 cm dish were allowed to grow for 36 h in complete DMEM. The medium was aspirated and replaced with serum-free DMEM and the cells were treated with 0 - 5 mM ropivacaine for 48 h. DNA was extracted as described previously 30 and analyzed by electrophoresis on a 1.6 % agarose gel.
Ioslation of protein extracts and Western blot analysis
Untreated or ropivacaine-treated HaCaT, HaCaTNeo, HaCaTBcl-2/2 or HaCaTApoJ/2 cells were washed with ice-cold PBS, resuspended in RIPA buffer containing protease inhibitors and incubated on ice for 20 min. Supernatants were clarified by centrifugation at 4°C and protein concentrations were determined with BioRad protein assay reagent. 30-60 μg of protein was analyzed by SDS-polyacrylamide gel electrophoresis followed by immunoblotting. The blots were blocked in 5% powdered milk in PBS containing 0.1% Tween-20 and probed with primary antibodies: anti-clusterin/ApoJ, anti-Bcl-2 and anti-caspase-3 (Santa Cruz Biotechnology, USA), anti-PARP (CII-10) (BD Transduction Laboratories, USA) and anti-actin (Sigma Chemical Co., Germany) followed by horseradish peroxidase (HRP)-conjugated secondary antibodies (Santa Cruz Biotechnology, USA). Antibody binding was detected using the Super Signal West Pico Chemiluminescence detection system (Pierce, USA).
RESULTS
Ropivacaine inhibited the proliferation and induced apoptosis of HaCaT cells
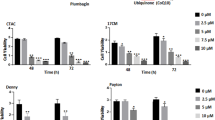
HaCaT cells were treated with increasing concentrations of ropivacaine (0 - 5 mM) in the absence of serum for 24 or 48 h in order to study its effects on cell proliferation. As shown in Fig. 1A, ropivacaine induced growth inhibition in a dose- and time-dependent manner. The inhibitory effect of ropivacaine on HaCaT cell proliferation was marked by 24 h and became more pronounced after 48 h of incubation, producing a 50% reduction in cell proliferation at 2 mM (Fig. 1A).

The effects of ropivacaine on the proliferation and viability of HaCaT cells. (A) Cells were treated with 0 - 5 mM ropivacaine for 24 h or 48 h in the absence of serum and counted. Experiments were performed three times in duplicates. Data shown are the means ± SD (p < 0.05). (B) Cells were treated with 0 - 5.0 mM ropivacaine for 48 h in the absence of serum and cell viability was evaluated by trypan blue exclusion assay.
The inhibition of cell proliferation by ropivacaine prompted us to determine whether ropivacaine reduced cell viability. HaCaT cells were treated with 0 - 5 mM ropivacaine for 48 h in the absence of serum and were subjected to trypan blue exclusion assay. We found that cell viability was reduced by ropivacaine in a dose-dependent manner. Exposure of HaCaT cells to 2 mM ropivacaine caused a cytotoxic effect with a reduction in cell viability by about 50% (Fig. 1B). HaCaT cells treated with ropivacaine displayed significantly morphological changes with increasing ropivacaine concentrations, as observed under a phase contrast microscope, characteristic of an apoptotic phenotype (data not shown).
Apoptosis has been characterized biochemically by the cleavage of DNA into multiples of 180 bp nucleosomal fragments. To further study the induction of cell death by ropivacaine and in particular whether ropivacaine caused DNA fragmentation, DNA was isolated from floating and attached untreated HaCaT cells and from cells treated with 0 - 5 mM ropivacaine for 48 h in the absence of serum. Treatment of HaCaT cells with ropivacaine resulted in a dose-dependent induction of internucleosomal DNA fragmentation, producing a DNA ladder characteristic of cells undergoing apoptosis. Induction of apoptosis by ropivacaine in HaCaT cells was evident at 1 mM and became more pronounced at concentrations of equal or above 2 mM ropivacaine (Fig. 2A).

Dose-dependent induction of apoptosis by ropivacaine in HaCaT cells. Cells were treated with 0 - 5 mM ropivacaine for 48 h in the absence of serum. (A) DNA isolated from floating and attached cells was analyzed by electrophoresis on a 1.6% agarose gel as described under materials and methods. (B) Total proteins isolated from ropivacaine-treated HaCaT cells were analyzed by western blot for the expression of PARP. Indicated are the intact and cleavage forms for PARP.
Genetic and biochemical studies showed that caspase activation during apoptosis is a tightly regulated process, resulting in the activation of caspase-3 which cleaves a number of cellular proteins including poly (ADP-ribose) polymerase (PARP) 31. Ropivacaine, at concentrations above 1 mM, induced the cleavage of the known caspase-3 substrate, PARP, from the 115 kD polypeptide to 89 kD fragment (Fig. 2B).
Ropivacaine downregulated clusterin/ApoJ, a cell survival protein, in keratinocytes
One way for apoptotic stimuli to induce cell death is to downregulate the expression of anti-apoptotic genes. Hence we investigated the expression of clusterin/ApoJ, a cell survival protein with anti-apoptotic properties. Immunoblot analysis showed that ropivacaine-treatment of HaCaT cells for 48 h in the absence of serum resulted in a marked reduction in the protein levels of clusterin/ApoJ, particularly at doses of or above 2 mM ropivacaine (Fig. 3), suggesting that a reduction in the expression of the anti-apoptotic protein clusterin/ApoJ may get involved in the drug induced apoptosis of HaCaT cells.

Ropivacaine downregulated the expression of clusterin/ApoJ in HaCaT cells. Total cell lysates isolated from untreated cells and from cells treated with 0 - 5 mM ropivacaine for 48 h in the absence of serum were analyzed by western blot for the expression of clusterin/ApoJ or actin. Indicated are the intracellular form (60 kD) and the secreted form of clusterin/ApoJ (75-80 kD) appearing as a doublet of ∼36-40 kD, representing the α- and β-chains of the protein, under reduced conditions. Exposure time: clusterin/ApoJ, 10 min; Actin, 1 min.
Generation and characterization of human clusterin/ApoJ-expressing HaCaT cells
To further analyze the role of clusterin/ApoJ and have it compared with the well-known anti-apoptotic effects of Bcl-2 in ropivacaine-mediated apoptosis, we generated stable HaCaTNeo, HaCaTBcl-2 and HaCaTApoJ transfectants. Several G418-resistant clones were isolated and characterized for the expression of clusterin/ApoJ or Bcl-2 by immunoblotting (Fig. 4). Fig. 4A shows the expression of clusterin/ApoJ in HaCaT cell clone 2 (referred to as HacaTApoJ/2 thereafter). Transfected HaCaT cells expressed higher levels of clusterin/ApoJ than their untransfected or neo-transfected control counterparts (Fig. 4A). In agreement with previous studies on the various forms of ApoJ and their processing, a diffuse doublet of ∼36-40 kD band that corresponds to α- and β-chains of the secreted 75-80 kD clusterin/ApoJ was detected as the cleavage product at the Asp227-Ser228 residues after the reduction of the protein (sClu). And a 60 kD band that corresponds to the cytoplasmic form of clusterin/Apo J (cClu) which is not reduced still retained inside the cells [18, 28]. Similarly, human clusterin/ApoJ was also overexpressed in HaCaTApoJ clone 2. As shown in Fig. 4B, whereas no Bcl-2 protein expression was detected in HaCaT or in HaCaTNeo cells, Bcl-2-transfected HaCaT cell clone 2 (referred to as HaCaTBcl-2/2 thereafter) expressed the 26 kD Bcl-2 protein at very high levels, for which it was subjected to subsequent analysis.

Expression of human clusterin/ApoJ and Bcl-2 in transfected HaCaT cells. HaCaT cells were transfected with pcDNA3.1B, pcDNA3.1/hApoJ or pcDNA3/hBcl-2, carrying the human clusterin/ApoJ or Bcl-2 cDNA, respectively, and selected in G418 to generate stable transfectants. Colonies were then isolated and subjected to western blot analysis for the expression of: (A) clusterin/ApoJ or (B) Bcl-2. The blots showed the expression of the indicated proteins in representative clones. Exposure time: Clusterin/ApoJ, 3 min; Bcl-2, 1 min. cClu, intracellular clusterin/ApoJ; sClu, secreted form of clusterin/ApoJ.
Clusterin/ApoJ or Bcl-2 decreased the sensitivity of HaCaT cells to ropivacaine
To investigate whether clusterin/ApoJ or Bcl-2 altered the growth behaviour and inhibited apoptosis in response to ropivacaine, HaCaTNeo, HaCaTBcl-2/2 or HaCaTApoJ/2 were treated with increasing concentrations of ropivacaine (0 - 5 mM) for 48 h in the absence of serum and subjected to cell proliferation assay. Growth of HaCaTneo cells was significantly inhibited by ropivacaine in a dose-dependent manner, with a ∼50% reduction at 2 mM ropivacaine (Fig. 5A). However, this inhibitory effect of ropivacaine was less profound in HaCaTBcl-2/2 and HaCaTApoJ/2 cells. Cell viability of HaCaTNeo, HaCaTBcl-2/2 and HaCaTApoJ/2 following ropivacaine treatment for 48 h was determined by the trypan blue exclusion assay (Fig. 5B).

The effects of ropivacaine on the proliferation and viability of Neo-, Bcl-2- or clusterin/ApoJ-expressing HaCaT cells. (A) Cells were treated with 0 - 5 mM ropivacaine for 48 h in the absence of serum and counted. Data shown are the means ± SD of six determinations (P < 0.05). (B) Cells were treated with 0 - 5.0 mM ropivacaine for 48 h in the absence of serum and cell viability was evaluated by trypan blue exclusion assay.
Cell viability of HaCaTNeo was reduced by ropivacaine in a dose-dependent manner, following a similar pattern to their untransfected control counterparts (Fig. 1B). Exposure of HaCaTNeo cells to 2 mM ropivacaine produced a cytotoxic effect with a reduction in cell viability by about 50%. However, cell viability in Bcl-2 (HaCaTBcl-2/2) or clusterin/ApoJ (HaCaTApoJ/2) overexpressing cells maintained as ever when treated with different concentrations of ropivacaine (Fig. 5B). Thus, HaCaTBcl-2/2 and HaCaTApoJ/2 displayed a phenotype resistant to ropivacaine-mediated apoptosis.
To investigate whether clusterin/ApoJ or Bcl-2 altered the toxic effects of ropivacaine on HaCaT cells, DNA isolated from floating and attached untreated and ropivacaine-treated cells was analyzed by agarose gel electrophoresis (Fig. 6). Whereas HaCaTNeo cells underwent apoptosis following ropivacaine-treatment, in a dose-dependent manner, similarly to untransfected HaCaT cells, aberrant overexpression of Bcl-2 or clusterin/ApoJ decreased the sensitivity of HaCaT cells to ropivacaine-induced DNA fragmentation (Fig. 6A).

Ectopic expression of Bcl-2 or clusterin/ApoJ suppressed ropivacaine-induced apoptosis of HaCaT cells. (A) DNA isolated from attached and floating untreated and ropivacaine-treated HaCaTNeo cells, HaCaTBcl-2/2 or HaCaTApoJ/2 cells were analyzed by electrophoresis on a 1.6% agarose gel. (B) Proteolytic cleavage of PARP or reduction in procaspase-3 expression is prevented by overexpression of human Bcl-2 or clusterin/ApoJ. Neo-, Bcl-2 or clusterin/ApoJ-transfected HaCaT ells were treated with 0 - 5 mM ropivacaine for 48 h in the absence of serum and total proteins isolated from untreated and ropivacaine-treated cells were analyzed by western blot for the expression of PARP or procaspase-3. Indicated are the intact and cleavage forms for PARP (115 and 89 kD); pro-Cas3, procaspase-3.
Given that cleavage of poly (ADP-ribose) polymerase (PARP) is a hallmark of apoptosis, proteins extracted from floating and attached untreated and ropivacaine-treated HaCaTneo, HaCaTBcl-2/2 or HaCaTApoJ/2 cells were subjected to immunoblotting for the expression of PARP (Fig. 6B). While HaCaTNeo cells exhibited cleavage of PARP, showing an intact 115 kD band and the cleaved 89 kD band in response to the treatment of 1 mM ropivacaine or at higher doses, clusterin/ApoJ- or Bcl-2-expressing HaCaT cells contained only the intact form of the enzyme (Fig. 6B). As PARP is a caspase-3 substrate, we also examined the expression of procaspase-3 in the same protein samples. It showed that with 1 mM ropivacaine or at higher doses, procaspase-3 expression was reduced in HaCaTneo but not in HaCaTBcl-2/2 or HaCaTApoJ/2 cells, correlating with PARP cleavage (Fig. 6B). The above data indicats that the overexpression of clusterin/ApoJ or Bcl-2 protected HaCaT cells from ropivacaine-mediated apoptosis.
DISCUSSION
In this study, firstly we found that ropivacaine could inhibit the proliferation of HaCaT cells in a dose- and time-dependent manner in the absence of serum. Ropivacaine was previously shown to inhibit the growth of various cell types including non-transformed cultured adult human fibroblasts, endothelial cells, keratinocyes 13 and colon adenocarcinoma cells 16, at concentrations ranging from 100 to 500 μM.
A second finding was that ropivacaine induced apoptosis of HaCaT cells cultured in serum free medium in a dose-dependent manner. The induction of cell death of HaCaT cells by ropivacaine was demonstrated by trypan blue exclusion assay, DNA fragmentation analysis and by the proteolytic cleavage of the caspase-3 substrate, PARP.
Thirdly, the present report demonstrated the reduction of clusterin/ApoJ protein levels in HaCaT cells following ropivacaine-treatment. Since clusterin/ApoJ is a cell survival gene with anti-apoptotic function, hence its down-regulation by ropivacaine may be a mechanism by which the local anesthetic induced apoptosis in HaCaT cells. To this point, we generated HaCaT cells overexpressing clusterin/ApoJ or Bcl-2 and compared the anti-apoptotic activities of these two proteins following ropivacaine-treatment of HaCaT cells. Ectopic overexpression of clusterin/ApoJ or Bcl-2 protected HaCaT cells from ropivacaine-induced apoptosis, as demonstrated by DNA fragmentation, the expression of PARP and the reduction in procaspase-3 expression. Collectively, these findings suggest that clusterin/ApoJ expression may play an important role in toxic effects induced by ropivacaine.
While ropivacaine was previously shown to decrease cell survival of primary rat fibroblasts and monkey kidney tumor cells demonstrated by trypan blue exclusion assay 6, our findings clearly demonstrated that ropivacaine could induce apoptosis in HaCaT cells, as evidenced by DNA fragmentation, PARP cleavage and procaspase-3 reduction. However, bupivacaine was shown to have a more potent cytotoxic effect than ropivacaine on the same cell types 6. Other local anesthetics appear to have similar effects on various cell types 9, 10, 11, 12, 13, 14. Prilocaine was found to induce apoptosis in mouse and human osteoblastic cells 11 and lidocaine was shown to inhibit the proliferation of normal human fibroblasts, endothelial cells and keratinocytes 12, while enhancing bleomycin A2 cytotoxicity and DNA strand breakage in murine leukemia cells and human head and neck carcinoma cells 13 and imparing corneal wound healing 14. Dibucaine was shown to induce apoptosis in a neuroblastoma cell line 9 and in HL-60 10 cells through activation of various caspases in conjunction with the release of cytochrome c from mitochondria induced by a processed product of Bid and depolarization of the mitochondrial membrane potential 10. Although at certain concentrations ropivacaine induced apoptosis of HaCaT cells, we do not know if this was due to Bid cleavage and cytochrome c release. The cleaved product of Bid results in translocation of Bid to mitochondria and insertion into the mitochondrial membrane where it inhibits the anti-apoptotic action of Bcl-2 and results in the release of cytochrome c with the subsequent activation of caspases. The fact that Bcl-2 completely abolished ropivacaine-induced apoptosis of HaCaT cells may imply a similar mechanism of that of dibucaine and ropivacaine in apoptosis.
Clusterin/ApoJ expression has been implicated in tumorigenesis 18, through its anti-apoptotic properties. Overexpression of clusterin/ApoJ cDNA into human cancer cell lines greatly improved their ability to resistant chemotherapy, through inhibition of apoptosis both in vitro and in vivo 23, 32, 33. Use of antisense oligonucleotides to inhibit endogenous clusterin/ApoJ expression showed that it conferred chemosensitivity to various cancer cells both in vitro and in vivo 25, 26, 27, 33, 34. Collectively, the data shows that increased expression of clusterin/ApoJ in cancer cells conferred cytoprotection.
Several studies showed that the induction of cell death by genotoxic agents may result in either up-regulation or downregulation of clusterin/ApoJ in the apoptotic cells. However, the mechanisms accounting for these differences still remain unknown. For example, oxidative stress of human diploid fibroblasts 22, 35 or C2-ceramide-treatment of pancreatic cells 36 induced anti-proliferative responses and up-regulated clusterin/apoJ perhaps as an attempt to survive. In contrast, apoptotic stimuli such as UVB irradiation, dexamethasone-treatment 37, chemotherapeutic agents 23, 38 or heat shock 39 induced apoptosis by markedly decreasing the expression of clusterin/ApoJ, in a dose- and time-dependent manner 23. The downregulation of clusterin/apoJ in keratinocytes undergoing ropivacaine-mediated apoptosis is in agreement with previous studies 40. In addition, it was previously shown that clusterin/ApoJ was induced by NF-κB 41, the major transcription factor involved in immune and inflammatory responses as well as in the regulation of cell growth and apoptosis 42. Given that ropivacaine possesses anti-inflammatory properties 3, 4, 5, 6 and induces antiproliferative responses 6, 12, 15, it is tempting to speculate that the effects of ropivacaine on HaCaT cells and on the expression of clusterin/ApoJ were most likely mediated through NF-κB. However this hypothesis requires verification. The downregulation of clusterin/apoJ by ropivacaine suggests that its administration together with a chemotherapeutic agent may, in addition to analgesic effects, enhance the sensitivity of tumor cells to cytotoxic chemotherapy, and may be used, at least in part, in the treatment of local tumors.
References
O'Keeffe NJ, Healy TE . The role of new anesthetic agents. Pharmacol Ther 1999; 84: 233–48.
Owen MD, Dean LS . Ropivacaine. Expert Opin Pharmacother 2000; 1: 325–36.
Martinsson T, Haegerstrand A, Dalsgaard CJ . Effects of ropivacaine on eicosanoid release from human granulocytes and endothelial cells in vitro. Inflamm Res 1997; 46: 398–403.
Martinsson T, Oda T, Fernvik E, et al . Ropivacaine inhibits leukocyte rolling, adhesion and CD11b/CD18 expression. J Pharmacol Exp Ther 1997; 283: 59–65.
Zhang XW, Thorlacious H . Inhibitory actions of ropivacaine on tumor necrosis factor-α induced leukocyte adhesion and tissue accumulation in vivo. Eur J Pharmacol 2000; 392: R1–3.
De Iuliis A, Zanatta L, Vincenti E, Galzigna L . Differences of ropivacaine and bupivacaine relevant to anti-inflammatory activity, platelet aggregation and antioxidant activity in vitro. Il Farmaco 2001; 56: 153–57.
Di Padova M, Barbieri R, Fanciulli M, Arcuri E, Floridi A . Effect of local anesthetic ropivacaine on the energy metabolism of Ehrlich ascites tumor cells. Oncol Res 1998; 10: 491–8.
Scutari G, Marian M, Bindoli A, et al. Mitochondrial effects of L-ropivacaine, a new local anesthetic. Biochem Pharmacol 1998; 56: 1633–7.
Kim M, Lee YS, Mathews HL, Wurster RD . Induction of apoptotic cell death in a neuroblastoma cell line by dibucaine. Exp Cell Res 1997; 231: 235–41.
Arita K, Utsami T, Kato A, et al. Mechanism of dibucaine-induced apoptosis in promyelocytic leukemia cells (HL-60). Biochem Pharmacol 2000; 60: 905–15.
Nakamura K, Kido H, Morimoto Y, et al. Prilocaine induces apoptosis in osteoblastic cells. Can J Anaesth 1999; 46: 476–82.
Martinsson T, Haegerstrand A, Dalsgaard, CJ . Ropivacaine and lidocaine inhibits proliferation of non-transformed cultured adult human fibroblasts, endothelial cells and keratinocytes. Agents Actions 1993; 40: 78–85.
Lazo JS, Braun D, Meandzija B, et al. Lidocaine potentiation of bleomycin A2 cytotoxicity and DNA strand breakage in L1210 and human A-253 cells. Cancer Res 1985; 45: 2103–9.
Bisla K, Tanelian DL . Concentration-dependent effects of lidocaine on corneal epithelial wound healing. Invest Opthalmol Vis Sci 1992; 33: 3029–33.
Martinsson T . Ropivacaine inhibits serum-induced proliferation of colon adenocarcinoma cells in vitro. J Pharmacol Exp Ther 1999; 288: 660–4.
Cory S, Adams JM . Regulation of the cellular life-or-death switch. Nat Rev Cancer 2002; 2: 647–56.
Jones SE, Jomary C . Clusterin. Int J Biochem Cell Biol 2002; 34: 427–31.
Trougakos IP, Gonos ES . Clusterin/apolipoprotein J in human aging and cancer. Int J Biochem Cell Biol 2002; 34: 1430–48.
Sensibar JA, Sutkowski DM, Raffo A, et al. Prevention of cell death induced by tumour necrosis factor-a in LNCaP cells by overexpression of sulfated glycoprotein-2 (clusterin). Cancer Res 1995; 55: 2431–7.
Sintich SM, Steinberg J, Kozlowski JM, et al. Cytotoxic sensitivity to tumor necrosis factor-alpha in PC3 and LNCaP prostatic cancer cells is regulated by extracellular levels of SGP-2 (clusterin). Prostate 1999; 39: 87–93.
Miyake H, Hara S, Zellweger T, Kamidono S, Gleave ME, Hara I . Acquisition of resistance to Fas-mediated apoptosis by overexpression of clusterin in human renal cell carcinoma cells. Mol Urol 2001; 5: 105–11.
Dumont P, Chainiaux F, Eliaers F, et al. Overexpression of apolipoprotein J in human fibroblasts protects against cytotoxicity and premature senescence induced by ethanol and tetr-butylhydroperoxide. Cell Stress Chap 2002; 7: 23–35.
Miyake H, Nelson C, Rennie PS, Gleave ME . Acquisition of chemoresistant phenotype by overexpression of the anti-apoptotic gene testosterone-repressed prostate message-2 in prostate cancer xenograft models. Cancer Res 2000; 60: 2547–54.
Viard I, Wehrli P, Jornot L, et al. Clusterin gene expression mediates resistance to apoptotic cell death induced by heat shock and oxidative stress. J Invest Dermatol 1999; 112: 290–6.
Miyake H, Chi KN, Gleave ME . Antisense TRPM-2 oligonucleotides chemosensitize human androgen-independent PC-3 prostate cancer cells both in vitro and in vivo. Clin Cancer Res 2000; 6: 1655–63.
Zellweger T, Miyake H, July LV, et al. Chemosensitization of human renal cell cancer using antisense oligonucleotides targeting the anti-apoptotic gene clusterin. Neoplasia 2001; 3: 360–7.
Miyake H, Hara I, Kamidono S, Gleave ME . Synergistic chemosensitization and inhibition of tumor growth and metastasis by the antisense oligonucleotide targeting clusterin gene in a human bladder cancer model. Clin Cancer Res 2001; 7: 4245–52.
Petropoulou C, Trougakos IP, Kolettas E, Toussaint O, Gonos ES . Clusterin/apolipoprotein J is a novel biomarker of cellular senescence that does not affect the proliferative capacity of human diploid fibroblasts. FEBS Lett 2001; 509: 287–97.
Odajima J, Matsumara I, Sonoyama J, et al. Full oncogenic activities of v-Src are mediated by multiple signaling pathways. Ras as an essential mediator for cell survival. J Biol Chem 2000; 275: 24096–105.
Sugiki H, Hozumi Y, Maeshima H, et al. C2-ceramide induces apoptosis in human squamous cell carcinoma cell line. Br J Dermatol 2000; 143: 1154–63.
Hengartner MO . The biochemistry of apoptosis. Nature 2000; 407: 770–6.
Miyake H, Nelson C, Rennie PS, Gleave ME . Testosterone-repressed prostate message-2 is an antiapoptotic gene involved in progression to androgen independence in prostate cancer. Cancer Res 2000; 60: 170–6.
Gleave ME, Miyake H, Zangemeister-Wittke U, Jansen B . Antisense therapy: current status in prostate cancer and other malignancies. Cancer Metastasis Rev 2002; 21: 79–92.
Trougakos IP, So A, Jansen B, Gleave ME, Gonos ES . Silencing expression of the clusterin/apolipoprotein j gene in human cancer cells using small interfering RNA induces spontaneous apoptosis, reduced growth ability, and cell sensitization to genotoxic and oxidative stress. Cancer Res 2004; 64: 1834–42.
Dumont P, Burton M, Chen QM, et al. Induction of replicative senescence biomarkers by sublethal oxidative stresses in normal human fibroblasts. Free Rad Biol Med 200; 28: 361–73.
Calvo EL, Mallo GV, Fiedler F, et al. Clusterin overexpression in rat pancreas during the acute phase of pancreatitis and pancreatic development. Eur J Biochem 1998; 254: 282–9.
French LE, Wohlwend A, Sappino AP, Tschopp J, Schiffereli JA . Human clusterin gene expression is confined to surviving cells during in vitro programmed cell death. J Clin Invest 1994; 93: 877–84.
Miyake H, Hara I, Kamidono S, Gleave ME, Eto H . Resistance to cytotoxic chemotherapy-induced apoptosis in human prostate cancer cells is associated with intracellular clusterin expression. Oncol Rep 2003; 10: 469–73.
Wu AJ, Park II, Zhaung L, Lee C . Response to a lethal dose of heat shock by a transient up-regulation of clusterin expression followed by down-regulation and apoptosis in prostate and bladder cancer cells. Prostate 2002; 53: 277–85.
Koch-Brandt C, Morgans C . Clusterin: a role in cell survival in the face of apoptosis? Prog Mol Cell Biol 1996; 16: 130–49.
Li X, Massa PE, Hanidu A, et al. IKKα, IKKβ and NEMO/IKKγ are each required for the NF-κB mediated inflammatory response program. J Biol Chem 2002; 277: 45129–40.
Li Q, Verma IM . NF-κB regulation in the immune system. Nature Rev Immunol 2002; 2: 725–34.
Acknowledgements
This work was supported in part by a grant from the European Society for Regional Anesthesia (ESRA). Work in ESG's laboratory has been supported by the following European Union full-costs grants: “Cellage” (QLK6-CT-2001-00616) and “Functionage” (QLK6-CT-2001-00310).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
KONTARGIRIS, E., KOLETTAS, E., VADALOUCA, A. et al. Ectopic expression of clusterin/apolipoprotein J or Bcl-2 decreases the sensitivity of HaCaT cells to toxic effects of ropivacaine. Cell Res 14, 415–422 (2004). https://doi.org/10.1038/sj.cr.7290242
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/sj.cr.7290242
Keywords
This article is cited by
-
Regulation of psoriasis, colitis, and the intestinal microbiota by clusterin
Scientific Reports (2023)
-
Zinc Inhibits Apoptosis and Maintains NEP Downregulation, Induced by Ropivacaine, in HaCaT Cells
Biological Trace Element Research (2012)
