
ABSTRACT
A number of studies show that environmental stress conditions increase abscisic acid (ABA) and hydrogen peroxide (H2O2) levels in plant cells. Despite this central role of ABA in altering stomatal aperture by regulating guard cell ion transport, little is known concerning the relationship between ABA and H2O2 in signal transduction leading to stomatal movement. Epidermal strip bioassay illustrated that ABA-inhibited stomatal opening and ABA-induced stomatal closure were abolished partly by externally added catalase (CAT) or diphenylene iodonium (DPI), which are a H2O2 scavenger and a NADPH oxidase inhibitor respectively. In contrast, internally added CAT or DPI nearly completely or partly reversed ABA-induced closure in half-stoma. Consistent with these results, whole-cell patch-clamp analysis showed that intracellular application of CAT or DPI partly abolished ABA-inhibited inward K+ current across the plasma membrane of guard cells. H2O2 mimicked ABA to inhibit inward K+ current, an effect which was reversed by the addition of ascorbic acid (Vc) in patch clamping micropipettes. These results suggested that H2O2 mediated ABA-induced stomatal movement by targeting inward K+ channels at plasma membrane.
Similar content being viewed by others

INTRODUCTION
Under stress conditions, ABA and H2O2 are generated as widespread molecules in many biological systems1, 2. It has been widely confirmed that ABA regulates stomatal movement as a stress signal1, yet there remain considerable gaps in our knowledge and in our attempts to construct a detailed description of the events underlying signaling chains involved in the process of stomatal closing3. On the other hand, reactive oxygen species (ROS) appear to play a crucial role in physiological and pathological processes of plants. H2O2, in particular, has been previously implicated as a second messenger in the regulation of plant hypersensitive response4, 5 and plays an important intermediary role in ABA signal transduction pathway leading to the induction of Cat1 gene6. Intriguingly, Pei et al have proved that H2O2 is involved in ABA-induced stomatal movement via the activation of calcium channels using Arabidopsis plants7.
K+ plays a significantly role in plant cells. This role has been best studied in guard cells where transmembrane K+ fluxes regulate cell volume, stomatal aperture, and thus gas exchange across the leaf. Therefore, plasma membrane K+ channels and their control are of importance in understanding these physiological processes. Interestingly, many environmental stress factors regulate stomatal aperture through modulation of ion channel activity in guard cells3. Some of these stress factors including drought, high salt and light intensity are also the factors that elevated ABA and ROS (especially H2O2, O2-). Based on the facts that ABA and ROS can inhibit inward K+ current at plasma membrane in guard cells, contributing to speeding up stomatal closure and inhibiting stomatal opening3, 8, 9, 10, and induce increases in cytoplasmic free Ca2+7, 11 and cytosolic pH12, we speculate that the elevation of H2O2 level may be related to ABA in the regulation of stomatal movement. In the present study, we provide evidences that ABA can inhibit inward K+ current at plasma membrane via a pathway of H2O2 generation. These results suggest that studies of oxidative signal molecule may be helpful in the better understanding of ABA signaling.
MATERIALS AND METHODS
Plant material
Broad bean (Vicia faba L.) was grown in a greenhouse with a humidity of about 70%, a photon flux density of 0.20 to 0.30 mmol m-2 s-1 and an ambient temperature (day/night 25±2 °C /20±2°C). Immediately prior to each experiment, the epidermis was peeled carefully from the abaxial surface of the youngest and fully expanded leaves of 4-week-old plants, and cut into pieces of 5-mm length.
Epidermal strip bioassay
To study the inhibition of stomatal opening, freshly prepared abaxial epidermis was first incubated in CO2-free 50 mM KCl/10 mM Mes, pH 6.15 (Mes-KCl). After a 1 h incubation in the dark, the epidermis was then transferred to CO2-free Mes-KCl in the presence or absence of ABA (1.0 μM) with or without 100 U/μl CAT or 10 μM DPI for another 2 h under conditions promoting stomatal opening (at 22 °C to 25 °C, under a photon flux density of 0.20 to 0.30 mmol m−2 s−1). To study the promotion of stomatal closure, freshly prepared abaxial epidermis was first incubated in CO2-free Mes-KCl for 3 h under conditions promoting stomatal opening to open the stomata, and the epidermis was then transferred to CO2-free Mes-KCl in the presence or absence of ABA (1.0 μM) with or without CAT or DPI for another 2 h under conditions promoting stomatal opening. In all cases, stomatal apertures were examined at the end of the subsequent 2 h incubation under a light microscope.
Microinjection of stomatal guard cells
To study the effects of microinjected reagents on stomata behavior, stomata with apertures of 8 to 10 μm were chosen for microinjection. The procedure was performed under a TE300 (objective 40×0.60 Plan Fluor) inverted microscope (Nikon) with a micromanipulator system (188NE, Narishige Scientific Instruments) according to methods of Ma et al13, Perona et al 14 and Schwartz et al8 with some modifications. Briefly, micropipettes (tip diameter 0.5 μm) for injection were made of borosillicate glass capillaries (GD-1, Narishige Scientific Instruments) by a micropipette puller (PC-10, Narishige Scientific Instruments). Micropipetter tips were frontfilled with injection reagents by applying a negative pressure. Micropipettes contained buffer (50 mM KCl /10 mM Tris, pH 7.2) with treatment reagents. The microinjection tip was inserted no more than 3 μm into the cytoplasm. Five minutes after microinjection, micropipette tip was slowly removed from the cell and allowed to recover for 10 min or so. Guard cells that failed to recover normally or had any visible morphological changes should be discarded at this time. Experimental guard cells with stomata that recovered (or nearly recovered) to the same aperture as those on the rest of the epidermal strip and in which both the injected and uninjected partners of a single stoma exhibited the same increase in turgor were maintained under white light at 0.20 mmol m−2 s−1 for another 30min, and stomatal apertures were recorded and measured. To further confirm the cell viability, after each experiment, FDA staining experiments showed the experimental cells still had high viability.
Whole-cell recordings
Guard cell protoplasts were isolated as described before15. Guard cell protoplasts were placed in bath solution (10 mM K-glutamate, 1 mM CaCl2, 2 mM MgCl2, 10 mM Mes, pH 5.5, and osmolality at 460 mOsM adjusted with mannitol). Recording pipettes were made from borosilicate glass capillaries (Kimax-51, Klimble Glass, Vineland, NJ, USA) by using a vertical two-stage puller (model PC-10; Narishige) and fire-polished by a microforge (model MF-90; Narishige) before use. The pipette solution contained 100 mM K-glutamate, 1.1 mM EGTA, 1.1 mM Mg-ATP, 2 mM MgCl2, 10 mM Hepes, pH 7.2, and osmolality at 510 mOsM adjusted with mannitol. Whole-cell patch-clamp recordings were performed essentially as described previously10, 16. Seal resistance was between 1 and 4GW in all experiments. Cell capacitance was each measured using the capacity compensation device of the amplifier and was between 3.6 and 7.5 pF. Data were acquired 15 min after the formation of the whole-cell configuration.
After the whole-cell configuration was obtained, the membrane was clamped to -52 mV (holding potential). Whole-cell currents were measured in response to 3s voltage pulse from -190 to -10 mV in 20-mV steps, using an EPC-9 patch-clamp amplifier (HEKA Elektronik, Lambrecht, Germany). Whole-cell data were low-pass filtered with a cut-off frequency of 2.9 kHz and analyzed with the software PLUSE and PLUSEFIT (version 8.3).
Leak currents were subtracted before generating whole-cell current-voltage relations. Leak currents for each cell were defined from the first 1 to 3 data points obtained after the membrane potential beginning from the holding voltage to the test voltages. The mean values of time-activated whole-cell currents were determined as the average of samples obtained between 2.0 and 2.5 s (500 samples total) after imposition of the test voltage (i.e. when the current amplitude had reached a plateau). After leak currents were subtracted, the final whole-cell currents were expressed as currents per unit capacitance (pA pF-1) to account for variations in the cell surface area.
Chemicals
Cellulase RS was from Yakult Honsha (Tokyo, Japan). Pectolyase Y-23 was from Seishin Pharmaceutical (Tokyo, Japan). Catalase (Bovine liver) and Cellulysin were from CalBiochem (La. Jolla, CA, USA). ABA (±), diphenylene iodonium (DPI), and ascorbic acid (Vc), Mes, Hepes, Mg-ATP, BSA, EGTA, K-glutamate were from Sigma. Unless stated otherwise, other chemicals were of analytical grade from Chinese companies.
RESULTS
ABA-induced stomatal movement was reversed by CAT and DPI
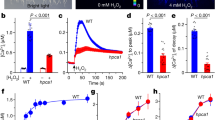
To gain insights into the mechanism of ABA induced-stomatal pore changes, we analyzed whether H2O2 is involved in ABA effects on stomatal movements, which may be similar to H2O2 effects in elicitor-induced defense responses. Vicia epidermal tissues were treated with 1 μM ABA in the presence of either 100 U/ml catalase (CAT) or 10 μM DPI, which can remove H2O2 and reduce the generation of H2O2 respectively17, 18, 19. As noted in Fig 1, both reagents reversed the ABA-inhibited stomatal opening (Fig 1A) or ABA-induced stomatal closure (Fig 1B), indicating that ABA affected stomatal behavior via a pathway involving H2O2 generation.

Effects of CAT and DPI on ABA-inhibited stomatal opening (A) and ABA-induced stomatal closure (B). Note that 100 U/ml CAT and 10 μM DPI, which reduce the level of H2O2, partly reversed the effects of ABA (1 μM) on stomatal movements. Experimental procedures are described in “Materials and Methods”. Values shown are the mean±SE (n=100) of three independent experiments.
To further confirm the effects of CAT and DPI on ABA-induced stomatal behavior, micoinjection was used to study the location of ABA action in intact guard cells. Fig 2 illustrated that microinjection of ABA into guard cells resulted in a significant (P < 0.05) reduction in stomatal half-aperture from 4.643±0.320 μm to 0.978±0.192 μm and co-microinjection of ABA and DPI resulted in a reduction in stomatal half-aperture from 4.769±0.287 μm to 2.910±0.417 μm (P < 0.05), whereas co-microinjection of ABA and CAT had no marked change in the half-aperture of the stomata, similar to the results following buffer microinjection. These findings suggest that H2O2 is a possible intermediate of the signal transduction pathway of ABA, and an ABA receptor site is also localized internally.

Effects of reagents or buffer microinjection on stomatal aperture in Vicia faba. One guard cell in a pair was injected. In each case, white bars (□) represent the half-apertures of the uninjected guard cells and black bars (▪) represent the half-aperture of the injected guard cells. Values shown are the mean±SE (n = 10 for each treatment).
ABA-induced inhibition of inward K + current was disrupted by CAT and DPI
After we observed the reversal effect of CAT and DPI on ABA-induced stomatal movements, we examined whether CAT and DPI regulate inward K+ current inhibited by ABA in guard cells using patch-clamp techniques. Fig 3A depicted a typical current recording obtained from patch clamp analysis of guard cell protoplast. This current was previously identified as K+ current by tail-current analysis 10, 20. 1 μM ABA in the bath solutions caused dramatic inhibition by approximately 76%, from-115pA pF−1 to -28pA pF−1 at -190 mV (Fig 3E). Addition of 100 U/ml CAT in the patch clamping micropipettes partly reversed ABA-induced inhibition of whole-cell inward K+ current (Fig 3D and Fig 3E). In a parallel experiment, addition of 10 μM DPI in the patch clamping micropipettes, which is a specific inhibition of NADPH oxidase contributing to H2O2 generation, also partly reversed ABA-induced inhibition of whole-cell inward K+ current similar to the effect of CAT (Fig 4D and Fig 4E). While application of CAT or DPI alone has no influence upon the inward K+ current.

Effects of CAT on ABA-induced inhibition of inward K+ current (Ikin) in Vicia guard cell protoplasts. Whole-cell patch-clamp recordings reveal the typical Ikin current (A), Ikin current with 1 μM ABA in the bath solution (B), Ikin current with 100 U/ml CAT in the pipette solution (C), and Ikin current with 1 μM ABA in the bath solution and 100 U/ml CAT in the pipette solution (D). Voltage protocols and the current/time scale bars for all whole-cell recordings are shown as in B. The current/voltage relations for various treatments are shown in E. The replicates for each treatment are 8 (A), 10 (B), 9 (C), and 10 (D), respectively.

Effects of DPI on ABA-induced inhibition of inward K+ current (Ikin) in Vicia guard cell protoplasts. Whole-cell patch-clamp recordings reveal the typical Ikin current (A), Ikin current with 1 μM ABA in the bath solution (B), Ikin current with 10 μM DPI in the pipette solution (C), and Ikin current with 1 M ABA in the bath solution and 10 μM DPI in the pipette solution (D). Voltage protocols and the current/time scale bars for all whole-cell recordings are shown as in B. The current/voltage relations for various treatments are shown in E. The replicates for each treatment are 6 (A), 7 (B), 10 (C), and 8 (D), respectively.
H 2 O 2 mimicked ABA to inhibit inward K + current
To further verify above findings, we directly added H2O2 in bath solutions to study whether H2O2 has the same effects on inward K+ current as ABA or not. As shown in Fig 5B and Fig 5E, 10-5 M H2O2 in the bath solutions significantly inhibited (67%; at -190 mV) whole-cell inward K+ current. Addition of 10 mM ascorbic acid (Vc), a widely used H2O2 scavenger21, in the micropipette, nearly completely reversed H2O2-induced inhibition of inward K+ current (Fig 5D and Fig 5E).

Effects of H2O2 on whole-cell inward K+ current (Ikin) in Vicia guard cell protoplasts. Whole-cell patch-clamp recordings reveal the typical Ikin current (A), Ikin current with 10−5 M H2O2 in the bath solution (B), Ikin current with 10 m M Vc in the pipette solution (C), and Ikin current with 10−5 M H2O2 in the bath solution and 10 m M Vc in the pipette solution (D). The current/voltage relations for various treatments are shown in E. Voltage protocols and the current/time scale bars for all whole-cell recordings are shown as in B. The replicates for each treatment are 10 (A), 8 (B), 11 (C), and 9 (D), respectively.
DISCUSSION
ABA-induced stomatal movement is one of the well-studied models of signaling systems, and some key signaling intermediates and effectors in ABA-induced stomatal closure have been identified, such as the increase of cytoplasmic Ca2+ ([Ca2+]i) and pH, the occurrence of protein phosphorylation events, and the change in ion flux22. However, the relationships between signaling intermediates remain unclear3. The present experiments showed that ABA affected guard cells via the generation of H2O2, targeting inward K+ channels, and that H2O2 mimicked ABA in the regulation of stomatal behavior. Epidermis bioassay showed that externally added CAT and DPI had similar effects on ABA-induced stomatal closure and opening (Fig 1A, B). In contrast, internally injected CAT completely abolished ABA-induced stomatal closure, while external CAT only partly abolished this process (compare Fig 2 with Fig 1B). A possible explanation might be that externally added CAT, which is remained in the apoplast, only partly removed H2O2 generated in guard cells after challenged with ABA. According to our recent findings that ABA-induced H2O2 might have several sources, at least two loci including chloroplasts and plasma membrane (NADPH oxidase) (to be published).
To date, regulation of guard cell ion transport by ABA, particular ABA inhibition of a guard cell inward K+ current is well documented23, but knowledge about up-regulation of the events and signaling chains remains very incomplete. Recent findings have illustrated that external and internal ABA can induce H2O2 generation in guard cells7, 24. Moreover, our previous work using voltage clamp has demonstrated that the stomata closure induced by externally applied H2O2 is due to the inhibition of K+ uptake and the evoking of K+ release through K+ channels on the plasma membrane of guard cells10. H2O2 is extremely sensitive to CAT and flavin-dependent enzymes including the mammalian NADPH oxidase, which are strongly inhibited by DPI 2, 18. Our patch-clamping experiments showed that intracellular application of CAT or DPI partly abolished ABA-inhibition of inward K+ current across the plasma membrane of guard cells (Fig 3, Fig 4). H2O2 mimicked ABA to inhibit inward K+ current, which effect was reversed by ascorbic acid (Vc) in the patch clamping micropipettes (Fig 5). However, treatments of the epidermis with CAT or DPI alone did not cause any changes of stomatal aperture (data not shown), which was consistent with the patch-clamping results. Additionally, based on recent findings that water stress-induced ABA accumulation is not affected either by free radicals, such as O2- and H2O2, or by oxidants such as KIO425, we think that H2O2 does not induce ABA generation in Vicia guard cells.
In addition, Cytosolic Ca2+ ([Ca2+]i) has been implicated as a signaling molecule in guard cell responses to oxidative signals11, 26 and has been reported to inhibit the inward K+ channels3. Recently a breakthrough in the direct recording of Ca2+ currents has been made7, 27 and Ca2+ channels and anion channels at the plasma membrane of stomatal guard cells are activated by hyperpolarization and abscisic acid7, 11, 28, 29. The increase of [Ca2+]i resulting from the activation of Ca2+ channels leading to Ca2+ influx is known to inactivate inward-rectifying K+ channels, biasing the plasma membrane for solute efflux, which in turn drives stomatal closure30. Up to now, the mechanism by which ABA leading to the activation and inactivation of specific ion channels and the relationships between H2O2 and other known messengers in ABA signaling are well worth studying.
References
Assmann SM, Shimazaki KI . The multisensory guard cell. Stomatal responses to blue light and abscisic acid. Plant Physiol 1999; 119:809–15.
Potikha TS, Collins CC, Johnson DI, Delmer DP, Levine A . The involvement of hydrogen peroxide in the differentiation of secondary walls in cotton fibers. Plant Physiol 1999; 119:849–58.
MacRobbie EAC . Signal transduction and ion channels in guard cells. Phil Trans R Soc Lond B 1998; 353:1475–88.
Mehdy MC . Active oxygen species in plant defense against pathogens. Plant Physiol 1994; 105:467–72.
Low PS, Merida JR . The oxidative burst in plant defense-function and signal transduction. Plant Physiol 1996; 96:533–42.
Guan L, Zhao J, Scandalios JG . Cis-elements and trans-factors that regulate expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response. Plant J 2000; 22:87–95.
Pei ZM, Murata Y, Benning G, Thomine S, Klusener B, Allen GJ, Grill E, Schroeder JL . Calcium channels activated by hydrogen peroxide mediate abscisic acid signaling in guard cells. Nature 2000; 406:731–34.
Schwartz A, Wu WH, Tucker EB, Assmann SM . Inhibition of inward K+ channels and stomatal response by abscisic acid: an intracllular locus of phytohormone action. Proc Natl Acad Sci USA 1994; 91:4019–23.
Torsethaugen G, Pell EJ, Assmann SM . Ozone inhibits guard cell K+ channels implicated in stomatal opening. Proc Natl Acad Sci USA 1999; 96:13577–82.
An GY, Song CP, Zhang X, Jing YC, Yang DM, Huang MJ, Wu CH, Zhou PA . Effect of hydrogen peroxide on stomatal movement and K+ channel on plasma membrane in Vicia faba guard cell. Acta Phytophysiol Sin 2000; 26:458–63.
Allen GJ, Chu SP, Schumacher K, Shimazaki CT, Vafeados D, Kemper A, Hawke SD, Tallman G, Tsien RY, Harper JF, Chory J, Schroeder JI . Alternation of stimulus-specific guard cell calcium oscillations and stomatal closing in Arabidopsis det3 mutant. Science 2000; 289:2338–42.
Zhang X, Dong FC, Gao JF, Song CP . Hydrogen peroxide-induced changes in intracellular pH of guard cells precede stomatal closure. Cell Res 2001; 11(1):37–43.
Ma LG, Xu XD, Cui SJ, Sun DY . The presence of a heterotrimeric G protein and its role in signal transduction of extracellular calmodulin in pollen germination and tube growth. Plant Cell 1999; 11:1351–63.
Perona R, Dolfi F, Feramisco J, Lacal JC . Microinjection of macromolecules into mammalian cells in culture. In Lacal JC, Perona R and Feramisco J . eds. Microinjection. Birkauser Verlag Basel: Switzerland 1999:17–30.
An GY, Song CP, Huang MJ, Yang DM, Zhou PA, Wu CH . Method of recording ion channels of plasma membrane in stomatal guard cells by patch clamp. Plant Physiol Commun. 1998; 34:129–32.
Wang XQ, Wu WH, Assmann SM . Differential responses of abaxial and adaxial guard cells of broad bean to abscisic acid and calcium. Plant Physiol 1998; 118:1421–9.
Lee S, Choi H, Suh S, Doo I-S, Oh K-Y, Choi EJ, Taylor SAT, Low PS, Lee Y . Oligogalacturonic acid and chitosan reduce stomatal aperture by inducing the evolution of reaction oxygen species from guard cells of tomato and Commelina communis. Plant Physiol 1999; 121:147–52.
Alvarez ME, Pennell RI, Meijer PJ, Ishikawa A, Dixon RA, Lamb C . Reactive oxygen intermediates mediate a systemic signal network in the establishment of plant immunity. Cell 1998; 92:773–84.
Levine A, Tenhaken R, Dixon RA, Lamb C . H2O2 from the oxidative burst orchestrates the plant hypersensitive response. Cell 1994; 79:583–93.
Wu W, Assmann SM . Photosynthesis by guard cell chloroplasts of Vicia faba L: Effects of factors associated with stomatal movements. Plant Cell Physiol 1993; 34:1015–22.
Thordal-Christensen H, Zhang ZZ, Wei YD, Collinge DB . Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J 1997; 11:1187–94.
Schroeder JI, Kwak JM, Allen GJ . Guard cell abscisic acid signaling and engineering drought hardiness in plants. Nature 2001; 410:327–30.
Sutton F, Paul SS, Wang XQ, Assmann SM . Distinct abscisic acid signaling pathways for modulation of guard cell versus mesophyll cell potassium channels revealed by expression studies in Xenopus laevis oocytes. Plant Physiol 2000; 124:223–30.
Zhang X, Zhang L, An GY, Gao JF, Song CP . Studies on ABA-induced H2O2 in Vicia guard cells by means of confocal laser scanning microscopy. Act Biol Exp Sin 2001; 34(1):71–6.
Jia W, Zhang J . Water stress-induced abscisic acid accumulation in relation to reducing agents and sulfhydryl modifiers in maize plant. Plant Cell and Environ. 2000; 23:1389–95.
McAinsh MR, Clayton H, Mansfield TA, Hetherington AM . Changes in stomatal behavior and cytosolic free calcium in response to oxidative stress. Plant Physiol 1996; 111:1031–42.
Hamilton D, Hills A, Kohler B, Blatt MR . Ca2+ channels at the plasma membrane of stomatal guard cells are activated by hyperpolarization and abscisic acid. Proc Natl Acad Sci USA 2000; 97:4967–72.
Pei ZM, Ghassemian M, Kwak CM, McCourt P, Schroeder JI . Role of farnesyltransferase in ABA regulation of guard cell anion channels and plant water loss. Science 1998; 282:287–90.
Li J, Wang XQ, Watson MB, Assmann SM . Regulation of abscisic acid-induced stomatal closure and anion channels by guard cell AAPK kinase. Science 2000; 287:300–3.
Blatt MR, Grabov A . Signaling gates in abscisic acid-mediated control of guard cell ion channels. Physiol Plant 1997; 100:481–90.
Acknowledgements
This work was supported by National Natural Science Foundation of China (No. 39870372), State Key Basic Research and Development Project (No. G1999011700), and Open Foundation of State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau (No. 10501-93).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
ZHANG, X., MIAO, Y., AN, G. et al. K+ channels inhibited by hydrogen peroxide mediate abscisic acid signaling in Vicia guard cells. Cell Res 11, 195–202 (2001). https://doi.org/10.1038/sj.cr.7290086
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/sj.cr.7290086
Keywords
This article is cited by
-
ATHK1 acts downstream of hydrogen peroxide to mediate ABA signaling through regulation of calcium channel activity in Arabidopsis guard cells
Chinese Science Bulletin (2013)
-
Compound stress response in stomatal closure: a mathematical model of ABA and ethylene interaction in guard cells
BMC Systems Biology (2012)
-
Brassica carinata CIL1 mediates extracellular ROS production during auxin- and ABA-regulated lateral root development
Journal of Plant Biology (2012)
-
The involvement of a P38-like MAP kinase in ABA-induced and H2O2-mediated stomatal closure in Vicia faba L.
Plant Cell Reports (2008)
