
Abstract
The beta-adrenergic receptors (ADRBs) are cell surface receptors that play central roles in the sympathetic nervous system. Pharmacological targeting of two of these receptors, ADRB1 and ADRB2, represents a widely used therapeutic approach for common and important diseases including asthma, hypertension and heart failure. Genetic variation in both ADRB1 and ADRB2 has been linked to both in vitro and clinical disease phenotypes. More recently, interest has shifted to studies that explore potential interaction between variation in ADRBs and medications directed at these important receptors. This paper reviews the current state of knowledge and understanding of ADRB genetic variation and explores the likely direction of future studies in this area.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 6 print issues and online access
$259.00 per year
only $43.17 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others
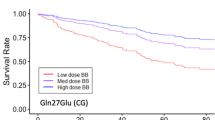

References
Busse W, Banks-Schlegel S, Noel P, Ortega H, Taggart V, Elias J . Future research directions in asthma: an NHLBI Working Group report. Am J Respir Crit Care Med 2004; 170: 683–690.
Palmer LJ, Burton PR, James AL, Musk AW, Cookson WO . Familial aggregation and heritability of asthma-associated quantitative traits in a population-based sample of nuclear families. Eur J Hum Genet 2000; 8: 853–860.
Palmer LJ, Cookson WO . Genomic approaches to understanding asthma. Genome Res 2000; 10: 1280–1287.
Palmer LJ, Silverman ES, Weiss ST, Drazen JM . Pharmacogenetics of asthma. Am J Respir Crit Care Med 2002; 165: 861–866.
Fuster V, Wayne A, O'Rourke R (eds). Hurst's The Heart, 11th edn McGraw-Hill: New York, 2004.
Port JD, Bristow MR . Altered beta-adrenergic receptor gene regulation and signaling in chronic heart failure. J Mol Cell Cardiol 2001; 33: 887–905.
Bristow MR . Changes in myocardial and vascular receptors in heart failure. J Am Coll Cardiol 1993; 22(4 Suppl A): 61A–71A.
Esler M, Kaye D, Lambert G, Esler D, Jennings G . Adrenergic nervous system in heart failure. Am J Cardiol 1997; 80: 7L–14L.
Tonolo G, Melis MG, Secchi G, Atzeni MM, Angius MF, Carboni A et al. Association of Trp64Arg beta 3-adrenergic-receptor gene polymorphism with essential hypertension in the Sardinian population. J Hypertens 1999; 17: 33–38.
Vendrell J, Gutierrez C, Broch M, Fernandez-Real JM, Aguilar C, Richart C . Beta 3-adrenoreceptor gene polymorphism and leptin. Lack of relationship in type 2 diabetic patients. Clin Endocrinol (Oxf) 1998; 49: 679–683.
Weiser M, Frishman WH, Michaelson MD, Abdeen MA . The pharmacologic approach to the treatment of obesity. J Clin Pharmacol 1997; 37: 453–473.
Gauthier C, Langin D, Balligand JL . Beta3-adrenoceptors in the cardiovascular system. Trends Pharmacol Sci 2000; 21: 426–431.
Gauthier C, Leblais V, Moniotte S, Langin D, Balligand JL . The negative inotropic action of catecholamines: role of beta3-adrenoceptors. Can J Physiol Pharmacol 2000; 78: 681–690.
Taylor MR, Bristow MR . The emerging pharmacogenomics of the beta-adrenergic receptors. Congest Heart Fail 2004; 10: 281–288.
Lelias JM, Kaghad M, Rodriguez M, Chalon P, Bonnin J, Dupre I et al. Molecular cloning of a human beta 3-adrenergic receptor cDNA. FEBS Lett 1993; 324: 127–130.
Granneman JG, Lahners KN, Rao DD . Rodent and human beta 3-adrenergic receptor genes contain an intron within the protein-coding block. Mol Pharmacol 1992; 42: 964–970.
van Spronsen A, Nahmias C, Krief S, Briend-Sutren MM, Strosberg AD, Emorine LJ . The promoter and intron/exon structure of the human and mouse beta 3-adrenergic-receptor genes. Eur J Biochem 1993; 213: 1117–1124.
Johnson JA, Terra SG . beta-Adrenergic receptor polymorphisms: cardiovascular disease associations and pharmacogenetics. Pharm Res 2002; 19: 1779–1787.
Farfel Z, Bourne HR, Iiri T . The expanding spectrum of G protein diseases. N Engl J Med 1999; 340: 1012–1020.
Moore JD, Mason DA, Green SA, Hsu J, Liggett SB . Racial differences in the frequencies of cardiac beta(1)-adrenergic receptor polymorphisms: analysis of c145A>G and c1165G>C. Hum Mutat 1999; 14: 271.
Podlowski S, Wenzel K, Luther HP, Muller J, Bramlage P, Baumann G et al. Beta1-adrenoceptor gene variations: a role in idiopathic dilated cardiomyopathy? J Mol Med 2000; 78: 87–93.
Borjesson M, Magnusson Y, Hjalmarson A, Andersson B . A novel polymorphism in the gene coding for the beta(1)-adrenergic receptor associated with survival in patients with heart failure. Eur Heart J 2000; 21: 1853–1858.
Bengtsson K, Orho-Melander M, Melander O, Lindblad U, Ranstam J, Rastam L et al. Beta(2)-adrenergic receptor gene variation and hypertension in subjects with type 2 diabetes. Hypertension 2001; 37: 1303–1308.
Bengtsson K, Melander O, Orho-Melander M, Lindblad U, Ranstam J, Rastam L et al. Polymorphism in the beta(1)-adrenergic receptor gene and hypertension. Circulation 2001; 104: 187–190.
Tesson F, Charron P, Peuchmaurd M, Nicaud V, Cambien F, Tiret L et al. Characterization of a unique genetic variant in the beta1-adrenoceptor gene and evaluation of its role in idiopathic dilated cardiomyopathy. CARDIGENE Group. J Mol Cell Cardiol 1999; 31: 1025–1032.
Yamada K, Ishiyama-Shigemoto S, Ichikawa F, Yuan X, Koyanagi A, Koyama W et al. Polymorphism in the 5′-leader cistron of the beta2-adrenergic receptor gene associated with obesity and type 2 diabetes. J Clin Endocrinol Metab 1999; 84: 1754–1757.
Small KM, Wagoner LE, Levin AM, Kardia SL, Liggett SB . Synergistic polymorphisms of beta1- and alpha2C-adrenergic receptors and. N Engl J Med 2002; 347: 1135–1142.
Reihsaus E, Innis M, MacIntyre N, Liggett SB . Mutations in the gene encoding for the beta 2-adrenergic receptor in normal and asthmatic subjects. Am J Respir Cell Mol Biol 1993; 8: 334–339.
Large V, Hellstrom L, Reynisdottir S, Lonnqvist F, Eriksson P, Lannfelt L et al. Human beta-2 adrenoceptor gene polymorphisms are highly frequent in obesity and associate with altered adipocyte beta-2 adrenoceptor function. J Clin Invest 1997; 100: 3005–3013.
Buscher R, Eilmes KJ, Grasemann H, Torres B, Knauer N, Sroka K et al. beta2 adrenoceptor gene polymorphisms in cystic fibrosis lung disease. Pharmacogenetics 2002; 12: 347–353.
Herrmann SM, Nicaud V, Tiret L, Evans A, Kee F, Ruidavets JB et al. Polymorphisms of the beta2-adrenoceptor (ADRB2) gene and essential hypertension: the ECTIM and PEGASE studies. J Hypertens 2002; 20: 229–235.
Ulbrecht M, Hergeth MT, Wjst M, Heinrich J, Bickeboller H, Wichmann HE et al. Association of beta(2)-adrenoreceptor variants with bronchial hyperresponsiveness. Am J Respir Crit Care Med 2000; 161(2 Part 1): 469–474.
Wood AJ . Variability in beta-adrenergic receptor response in the vasculature: role of receptor polymorphism. J Allergy Clin Immunol 2002; 110(6 Suppl): S318–S321.
Xie HG, Stein CM, Kim RB, Xiao ZS, He N, Zhou HH et al. Frequency of functionally important beta-2 adrenoceptor polymorphisms varies markedly among African-American, Caucasian and Chinese individuals. Pharmacogenetics 1999; 9: 511–516.
Liggett SB . beta(2)-adrenergic receptor pharmacogenetics. Am J Respir Crit Care Med 2000; 161(3 Part 2): S197–S201.
Liggett SB . Pharmacogenetics of beta-1- and beta-2-adrenergic receptors. Pharmacology 2000; 61: 167–173.
Liggett SB, Wagoner LE, Craft LL, Hornung RW, Hoit BD, McIntosh TC et al. The Ile164 beta2-adrenergic receptor polymorphism adversely affects the outcome of congestive heart failure. J Clin Invest 1998; 102: 1534–1539.
McGraw DW, Forbes SL, Kramer LA, Liggett SB . Polymorphisms of the 5′ leader cistron of the human beta2-adrenergic receptor regulate receptor expression. J Clin Invest 1998; 102: 1927–1932.
Fujisawa T, Ikegami H, Yamato E, Hamada Y, Kamide K, Rakugi H et al. Trp64Arg mutation of beta3-adrenergic receptor in essential hypertension: insulin resistance and the adrenergic system. Am J Hypertens 1997; 10: 101–105.
Hein L . Physiological significance of beta-adrenergic receptor polymorphisms: in-vivo or in-vitro veritas? Pharmacogenetics 2001; 11: 187–189.
Mason DA, Moore JD, Green SA, Liggett SB . A gain-of-function polymorphism in a G-protein coupling domain of the human beta1-adrenergic receptor. J Biol Chem 1999; 274: 12670–12674.
Maqbool A, Hall AS, Ball SG, Balmforth AJ . Common polymorphisms of beta1-adrenoceptor: identification and rapid screening assay. Lancet 1999; 353: 897.
dbSNP database.
Drysdale CM, McGraw DW, Stack CB, Stephens JC, Judson RS, Nandabalan K et al. Complex promoter and coding region beta 2-adrenergic receptor haplotypes alter receptor expression and predict in vivo responsiveness. Proc Natl Acad Sci U S A 2000; 97: 10483–10488.
Chamberlain PD, Jennings KH, Paul F, Cordell J, Berry A, Holmes SD et al. The tissue distribution of the human beta3-adrenoceptor studied using a monoclonal antibody: direct evidence of the beta3-adrenoceptor in human adipose tissue, atrium and skeletal muscle. Int J Obes Relat Metab Disord 1999; 23: 1057–1065.
Perfetti R, Hui H, Chamie K, Binder S, Seibert M, McLenithan J et al. Pancreatic beta-cells expressing the Arg64 variant of the beta(3)-adrenergic receptor exhibit abnormal insulin secretory activity. J Mol Endocrinol 2001; 27: 133–144.
Zhang C, Williams MA, Edwards KL, Austin MA . Trp64Arg polymorphism of the beta3-adrenergic receptor gene, pre-pregnancy obesity and risk of pre-eclampsia. J Matern Fetal Neonatal Med 2005; 17: 19–28.
Bachman ES, Dhillon H, Zhang CY, Cinti S, Bianco AC, Kobilka BK et al. betaAR signaling required for diet-induced thermogenesis and obesity resistance. Science 2002; 297: 843–845.
Moniotte S, Kobzik L, Feron O, Trochu JN, Gauthier C, Balligand JL . Upregulation of beta(3)-adrenoceptors and altered contractile response to inotropic amines in human failing myocardium. Circulation 2001; 103: 1649–1655.
Emorine L, Blin N, Strosberg AD . The human beta 3-adrenoceptor: the search for a physiological function. Trends Pharmacol Sci 1994; 15: 3–7.
Langin D, Tavernier G, Lafontan M . Regulation of beta 3-adrenoceptor expression in white fat cells. Fundam Clin Pharmacol 1995; 9: 97–106.
Allison DB, Heo M, Faith MS, Pietrobelli A . Meta-analysis of the association of the Trp64Arg polymorphism in the beta3 adrenergic receptor with body mass index. Int J Obes Relat Metab Disord 1998; 22: 559–566.
Strosberg AD . Association of beta 3-adrenoceptor polymorphism with obesity and diabetes: current status. Trends Pharmacol Sci 1997; 18: 449–454.
Walston J, Silver K, Bogardus C, Knowler WC, Celi FS, Austin S et al. Time of onset of non-insulin-dependent diabetes mellitus and genetic variation in the beta 3-adrenergic-receptor gene. N Engl J Med 1995; 333: 343–347.
Garcia-Rubi E, Calles-Escandon J . Insulin resistance and type 2 diabetes mellitus: its relationship with the beta 3-adrenergic receptor. Arch Med Res 1999; 30: 459–464.
Arner P, Hoffstedt J . Adrenoceptor genes in human obesity. J Intern Med 1999; 245: 667–672.
Leineweber K, Buscher R, Bruck H, Brodde OE . Beta-adrenoceptor polymorphisms. Naunyn Schmiedebergs Arch Pharmacol 2004; 369: 1–22.
Rathz DA, Brown KM, Kramer LA, Liggett SB . Amino acid 49 polymorphisms of the human beta1-adrenergic receptor affect agonist-promoted trafficking. J Cardiovasc Pharmacol 2002; 39: 155–160.
Levin MC, Marullo S, Muntaner O, Andersson B, Magnusson Y . The myocardium-protective Gly-49 variant of the beta 1-adrenergic receptor. J Biol Chem 2002; 277: 30429–30435.
Green SA, Turki J, Innis M, Liggett SB . Amino-terminal polymorphisms of the human beta 2-adrenergic receptor impart distinct agonist-promoted regulatory properties. Biochemistry 1994; 33: 9414–9419.
Dishy V, Sofowora GG, Xie HG, Kim RB, Byrne DW, Stein CM et al. The effect of common polymorphisms of the beta2-adrenergic receptor on agonist-mediated vascular desensitization. N Engl J Med 2001; 345: 1030–1035.
Green SA, Cole G, Jacinto M, Innis M, Liggett SB . A polymorphism of the human beta 2-adrenergic receptor within the fourth transmembrane domain alters ligand binding and functional properties of the receptor. J Biol Chem 1993; 268: 23116–23121.
Turki J, Lorenz JN, Green SA, Donnelly ET, Jacinto M, Liggett SB . Myocardial signaling defects and impaired cardiac function of a human beta 2-adrenergic receptor polymorphism expressed in transgenic mice. Proc Natl Acad Sci USA 1996; 93: 10483–10488.
Parola AL, Kobilka BK . The peptide product of a 5′ leader cistron in the beta 2 adrenergic receptor mRNA inhibits receptor synthesis. J Biol Chem 1994; 269: 4497–4505.
Johnatty SE, Abdellatif M, Shimmin L, Clark RB, Boerwinkle E . Beta2 adrenergic receptor 5′ haplotypes influence promoter activity. Br J Pharmacol 2002; 137: 1213–1216.
Ranade K, Jorgenson E, Sheu WH, Pei D, Hsiung CA, Chiang FT et al. A polymorphism in the beta1 adrenergic receptor is associated with resting heart rate. Am J Hum Genet 2002; 70: 935–942.
Humma LM, Puckett BJ, Richardson HE, Terra SG, Andrisin TE, Lejeune BL et al. Effects of beta1-adrenoceptor genetic polymorphisms on resting hemodynamics in patients undergoing diagnostic testing for ischemia. Am J Cardiol 2001; 88: 1034–1037.
Karlsson J, Lind L, Hallberg P, Michaelsson K, Kurland L, Kahan T et al. Beta1-adrenergic receptor gene polymorphisms and response to beta1-adrenergic receptor blockade in patients with essential hypertension. Clin Cardiol 2004; 27: 347–350.
Magnusson Y, Levin MC, Eggertsen R, Nystrom E, Mobini R, Schaufelberger M et al. Ser49Gly of beta1-adrenergic receptor is associated with effective beta-blocker dose in dilated cardiomyopathy. Clin Pharmacol Ther 2005; 78: 221–231.
Mialet Perez J, Rathz DA, Petrashevskaya NN, Hahn HS, Wagoner LE, Schwartz A et al. Beta 1-adrenergic receptor polymorphisms confer differential function and predisposition to heart failure. Nat Med 2003; 9: 1300–1305.
Wills-Karp M, Ewart SL . Time to draw breath: asthma-susceptibility genes are identified. Nat Rev Genet 2004; 5: 376–387.
Contopoulos-Ioannidis DG, Manoli EN, Ioannidis JP . Meta-analysis of the association of beta2-adrenergic receptor polymorphisms with asthma phenotypes. J Allergy Clin Immunol 2005; 115: 963–972.
Israel E, Chinchilli VM, Ford JG, Boushey HA, Cherniack R, Craig TJ et al. Use of regularly scheduled albuterol treatment in asthma: genotype-stratified, randomised, placebo-controlled cross-over trial. Lancet 2004; 364: 1505–1512.
Martinez FD, Graves PE, Baldini M, Solomon S, Erickson R . Association between genetic polymorphisms of the beta2-adrenoceptor and response to albuterol in children with and without a history of wheezing. J Clin Invest 1997; 100: 3184–3188.
Rosmond R, Ukkola O, Chagnon M, Bouchard C, Bjorntorp P . Polymorphisms of the beta2-adrenergic receptor gene (ADRB2) in relation to cardiovascular risk factors in men. J Intern Med 2000; 248: 239–244.
Bray MS, Krushkal J, Li L, Ferrell R, Kardia S, Sing CF et al. Positional genomic analysis identifies the beta(2)-adrenergic receptor gene as a susceptibility locus for human hypertension. Circulation 2000; 101: 2877–2882.
Bristow MR, Ginsburg R, Minobe W, Cubicciotti RS, Sageman WS, Lurie K et al. Decreased catecholamine sensitivity and beta-adrenergic-receptor density in failing human hearts. N Engl J Med 1982; 307: 205–211.
Bristow MR, Ginsburg R, Umans V, Fowler M, Minobe W, Rasmussen R et al. Beta 1- and beta 2-adrenergic-receptor subpopulations in nonfailing and failing human ventricular myocardium: coupling of both receptor subtypes to muscle contraction and selective beta 1-receptor down-regulation in heart failure. Circ Res 1986; 59: 297–309.
Brodde OE, Schuler S, Kretsch R, Brinkmann M, Borst HG, Hetzer R et al. Regional distribution of beta-adrenoceptors in the human heart: coexistence of functional beta 1- and beta 2-adrenoceptors in both atria and ventricles in severe congestive cardiomyopathy. J Cardiovasc Pharmacol 1986; 8: 1235–1242.
Wagoner LE, Craft LL, Singh B, Suresh DP, Zengel PW, McGuire N et al. Polymorphisms of the beta(2)-adrenergic receptor determine exercise capacity in patients with heart failure. Circ Res 2000; 86: 834–840.
Kaye DM, Smirk B, Finch S, Williams C, Esler MD . Interaction between cardiac sympathetic drive and heart rate in heart failure: modulation by adrenergic receptor genotype. J Am Coll Cardiol 2004; 44: 2008–2015.
Kaye DM, Smirk B, Williams C, Jennings G, Esler M, Holst D . Beta-adrenoceptor genotype influences the response to carvedilol in patients with congestive heart failure. Pharmacogenetics 2003; 13: 379–382.
Brain Jr N, Dominiczak AF . Pharmacogenomics in hypertension: present practicalities and future potential. J Hypertens 2005; 23: 1327–1329.
Mellen PB, Herrington DM . Pharmacogenomics of blood pressure response to antihypertensive treatment. J Hypertens 2005; 23: 1311–1325.
Sandilands A, Yeo G, Brown MJ, O'Shaughnessy KM . Functional responses of human beta1 adrenoceptors with defined haplotypes for the common 389R>G and 49S>G polymorphisms. Pharmacogenetics 2004; 14: 343–349.
Defoor J, Martens K, Zielinska D, Matthijs G, Van Nerum H, Schepers D et al. The CAREGENE study: polymorphisms of the {beta}1-adrenoceptor gene and aerobic power in coronary artery disease. Eur Heart J 2006; 27: 808–816.
Bruck H, Leineweber K, Park J, Weber M, Heusch G, Philipp T et al. Human beta2-adrenergic receptor gene haplotypes and venodilation in vivo. Clin Pharmacol Ther 2005; 78: 232–238.
Oostendorp J, Postma DS, Volders H, Jongepier H, Kauffman HF, Boezen HM et al. Differential desensitization of homozygous haplotypes of the beta2-adrenergic receptor in lymphocytes. Am J Respir Crit Care Med 2005; 172: 322–328.
Petrone A, Zavarella S, Iacobellis G, Zampetti S, Vania A, Di Pietro S et al. Association of beta2 adrenergic receptor polymorphisms and related haplotypes with triglyceride and LDL-cholesterol levels. Eur J Hum Genet 2006; 14: 94–100.
Zee RY, Cook NR, Reynolds R, Cheng S, Ridker PM . Haplotype analysis of the beta2 adrenergic receptor gene and risk of myocardial infarction in humans. Genetics 2005; 169: 1583–1587.
Muszkat M, Stein CM . Pharmacogenetics and response to beta-adrenergic receptor antagonists in heart failure. Clin Pharmacol Ther 2005; 77: 123–126.
Cascorbi I, Paul M, Kroemer HK . Pharmacogenomics of heart failure – focus on drug disposition and action. Cardiovasc Res 2004; 64: 32–39.
La Rosee K, Huntgeburth M, Rosenkranz S, Bohm M, Schnabel P . The Arg389Gly beta1-adrenoceptor gene polymorphism determines contractile response to catecholamines. Pharmacogenetics 2004; 14: 711–716.
Terra SG, Hamilton KK, Pauly DF, Lee CR, Patterson JH, Adams KF et al. Beta1-adrenergic receptor polymorphisms and left ventricular remodeling changes in response to beta-blocker therapy. Pharmacogenet Genomics 2005; 15: 227–234.
Terra SG, Pauly DF, Lee CR, Patterson JH, Adams KF, Schofield RS et al. beta-Adrenergic receptor polymorphisms and responses during titration of metoprolol controlled release/extended release in heart failure. Clin Pharmacol Ther 2005; 77: 127–137.
Litonjua AA, Silverman EK, Tantisira KG, Sparrow D, Sylvia JS, Weiss ST . Beta 2-adrenergic receptor polymorphisms and haplotypes are associated with airways hyperresponsiveness among nonsmoking men. Chest 2004; 126: 66–74.
Acknowledgements
Ms Sharon Bobel is acknowledged for her assistance with the preparation of the figures.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Taylor, M. Pharmacogenetics of the human beta-adrenergic receptors. Pharmacogenomics J 7, 29–37 (2007). https://doi.org/10.1038/sj.tpj.6500393
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.tpj.6500393
Keywords
This article is cited by
-
Dynamic FRET-FLIM based screening of signal transduction pathways
Scientific Reports (2021)
-
Activation of β2 adrenergic receptor signaling modulates inflammation: a target limiting the progression of kidney diseases
Archives of Pharmacal Research (2021)
-
Investigating changes in β-adrenergic gene expression (ADRB1 and ADRB2) in Takotsubo (stress) cardiomyopathy syndrome; a pilot study
Molecular Biology Reports (2021)
-
Conformational plasticity of ligand-bound and ternary GPCR complexes studied by 19F NMR of the β1-adrenergic receptor
Nature Communications (2020)
-
A Literature Review of Genetic Markers Conferring Impaired Response to Cardiovascular Drugs
American Journal of Cardiovascular Drugs (2018)
