
Abstract
Gastrointestinal stromal tumors (GISTs) may be caused by germline mutations of the KIT and platelet-derived growth factor receptor-α (PDGFRA) genes and treated by Imatinib mesylate (STI571) or other protein tyrosine kinase inhibitors. However, not all GISTs harbor these genetic defects and several do not respond to STI571 suggesting that other molecular mechanisms may be implicated in GIST pathogenesis. In a subset of patients with GISTs, the lesions are associated with paragangliomas; the condition is familial and transmitted as an autosomal-dominant trait. We investigated 11 patients with the dyad of ‘paraganglioma and gastric stromal sarcoma’; in eight (from seven unrelated families), the GISTs were caused by germline mutations of the genes encoding subunits B, C, or D (the SDHB, SDHC and SDHD genes, respectively). In this report, we present the molecular effects of these mutations on these genes and the clinical information on the patients. We conclude that succinate dehydrogenase deficiency may be the cause of a subgroup of GISTs and this offers a therapeutic target for GISTs that may not respond to STI571 and its analogs.
Similar content being viewed by others

Introduction
Gastrointestinal stromal tumors (GIST) may be sporadic or inherited in an autosomal-dominant manner either alone or as a component of a syndrome associated with other manifestations or predisposing to the development of multiple tumors.1 Recently, we described 12 young patients (seven male and five female with an average age of 23 years) from five unrelated families who had both GIST and paraganglioma; the tumors were inherited in an apparently autosomal-dominant manner, with incomplete penetrance.2 GISTs in these patients were multifocal and paragangliomas multicentric, supporting a genetic link between the two lesions. The condition has been referred to as the dyad of ‘paraganglioma and gastric stromal sarcoma’ or the ‘Carney–Stratakis syndrome’ or Carney–Stratakis dyad.3
Germline mutations of the genes encoding succinate dehydrogenase subunits B, C and D (SDHB, SDHC and SDHD) had been described in inherited paraganglioma and pheochromocytoma4, 5, 6, 7, 8 but not in familial GISTs, until patients with the dyad were studied.9 In this report, we describe the molecular and clinical effects of germline SDHB, SDHC and SDHD mutations in patients with the dyad and their tumors. Allelic losses around the SDHB and SDHC chromosomal loci were found in GISTs from these patients, pointing to a tumor-suppressor function of the succinate dehydrogenase subunits. These tumors did not exhibit the KIT or platelet-derived growth factor receptor-α (PDGFRA) gene mutations associated with GISTs and related lesions10, 11, 12 that are responsive to Imatinib mesylate (STI571) and its analogs.
Methods
Clinical studies and tissue samples
The Institutional Review Boards of the participating institutions have approved the DNA and other studies of this investigation. Families with the dyad have been described by Carney and Stratakis2 (Table 1). All patients had GIST or paraganglioma diagnosed by standard clinical and pathological criteria. Blood and tissue samples were collected from patients after informed consent, as described elsewhere.13 When possible, tissue was collected at surgery and processed for routine histopathology and immunohistochemistry following formalin fixation and paraffin embedment; additional fragments were frozen at −70°C. The antibodies for c-KIT, actin, desmin and vimentin were used at dilutions 1/1000, 1/1000, 1/200 and 1/500, respectively, and were obtained from DAKO (Carpinteria, CA, USA); the antibody for CD34 was used at a dilution of 1/100 and was obtained from BD Biosciences (San Jose, CA, USA); and the antibody for chromogranin A was diluted at 1/500 and was obtained from Chemicon (Temecula, CA, USA); for all the detection system was polymer based.
DNA was extracted from blood, frozen and archived tissue samples or cell lines using standard methods (Qiagen, Valencia, CA, USA). Tumor samples were microdissected from associated normal tissues.
Sequencing and allelic heterozygosity analysis
Mutation analysis for exons, exon–intron boundaries and flanking intronic regions was performed for the KIT, SDHA, SDHB, SDHC, SDHD and PDGFRA genes; polymerase chain reaction (PCR) was performed using Qiagen HotStarTaq kit (Qiagen) and primers that have been described elsewhere.5, 6, 8, 9, 10, 11, 12, 14, 15 An aliquot of the PCR product was purified with exonuclease I/shrimp alkaline phosphatase treatment (New England Biolabs, Beverly, MA, USA/USB Corporation, Cleveland, OH, USA). The purified amplicons were directly sequenced using Big-Dye Terminator v. 20 Cycle Sequencing kit (Applied Biosystems, Foster City, CA, USA) and analyzed on an ABI 3730 DNA analyzer (Applied Biosystems). Cell lines, blood cells or frozen tumor material were used for total RNA extraction, complementary DNA (cDNA) synthesis by reverse transcriptase PCR and sequencing, using appropriate transcribed sequence-based primers.
Tumor and DNA from other samples were also subjected to loss-of-heterozygosity (LOH) analysis using markers surrounding the genes in which mutations were identified (SDHB and SDHC, see below). Markers were selected using NCBI MapViewer35 (http://www.ncbi.nlm.nih.gov/mapview/); for each gene, at least 10 informative sequences were selected for heterozygosity studies, as described previously.13
For paternity and/or homozygosity testing (the latter for the twin brothers of family CTR02, see below), seven fluorescent polymorphic microsatellite markers located in three different chromosomes were used: they included D1S1595, D1S2624, D11S934, D11S968, D14S264, D14S283 and D14S990, which were amplified and run using standard procedures.
All sequence variants that were identified in the study patients were also searched for in more than 100 ethnically matched-control DNA samples. The mutations we report were not found in any of the control samples; numerous polymorphisms were identified in the samples of the study patients for all six genes that, however, were also present in the control samples (data not shown).
Search for microdeletions of SDHB, SDHC and SDHD and other loci
Tumor DNA was hybridized to Illumina (Illumina Inc., San Diego, CA, USA) HumanHap300-Duo Genotyping BeadChips, according to the manufacturer's protocol (www.illumina.com). Data analysis was performed using Illumina software BeadStudio (version 2) and R version 2.3 (R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna, Austria) (data not shown). For the single patient in whom SDHB was deleted (CTRS20; Table 1), confirmation of the hemizygous deletion was performed with semi-quantitative duplex PCR of each SDHB exon compared to the housekeeping GAPDH exon 8 gene sequence using a method that we have reported elsewhere.16 For the experiments reported here, 20 ng of genomic DNA was used as template for each semi-quantitative PCR reaction with primers flanking each SDHB exon and those of GAPDH exon 8 (Table 2) using Qiagen HotStartTaq kit (Qiagen) and 30 cycles with an annealing temperature of 55°C.
Results
Germline and tumor DNA studies
We accrued a total of nine families with the Carney–Stratakis dyad of ‘paraganglioma and gastric stromal sarcoma’ (Figures 1 and 2) from the United States, France and Italy. None had mutations of the PDGFRA or KIT genes (data not shown); however, seven probands had a mutation or a deletion in one of the three succinate dehydrogenase subunit-encoding genes (SDHB, SDHC or SDHD). The genomic DNA mutations have been published elsewhere.9 In this article, we report the clinical information of the patients and study the effects of these mutations on the transcribed message and/or tumors. There were no mutations in the coding sequence of the SDHA gene; a number of SDHA sequence variants were identified in the study patients but they were present in similar frequency in the control DNA (data not shown).

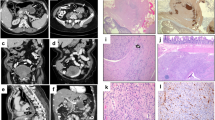
(a) Multiple tumors protrude from the anterior wall of stomach of patient CTRS35; two show surface bosselation. (b) Hematoxylin and eosin staining of one of the stomach GISTs from the CTRS35 patient; staining with (c) cKIT, (d) CD-34, (e) actin, (f) desmin and (g) vimentin.

(a) Retroperitoneal paraganglioma with dilated veins on its surface from patient CTRS35; (b) hematoxylin and eosin staining of the paraganglioma excised from patient CTRS35; and (c) staining with the neuroendocrine marker chromogranin A.
There were mutations in the SDHB gene in three families: in the first family, a 37-year-old male (CTRS35) had melena resulting from ulceration of a stomach GIST (Figure 1). During workup for metastatic disease, a non-functioning periaortic paraganglioma was found (Figure 2). The patient carried a relatively frequent germline mutation in intron 1 of the SDHB gene (IVS1+1 G>T or c.72+1G>T);5, 9, 14 cDNA analysis from his white blood cells revealed the heterozygous presence of an aberrant transcript that, when sequenced, indicated that the first part of intron 1 was transcribed (Figure 3) with the frameshift resulting in a stop codon in the middle of exon 2 and predicting a significantly truncated protein. The patient's GIST contained the mutant SDHB sequence only (Supplementary Figure 1), indicating loss of the normal SDHB allele. The patient's mother was operated for a pheochromocytoma at the age of 57; her DNA was not available.

cDNA studies (RT-PCR) of the CTRS35 SDHB IVS1+1 mutation (*): it caused intron 1 to be transcribed, resulting in a predicted truncated protein with a stop codon coded by a downstream sequence (318 bp) of intron 1.
In the second family (CTR02), monozygotic twin brothers were diagnosed with the Carney–Stratakis syndrome;17 one was operated at 12 years of age for a non-functioning paraganglioma of the organ of Zuckerkandl, and the other for a stomach GIST at age 13 years. Monozygosity was confirmed by polymorphic microsatellite markers: the twins shared identical haplotypes (Table 3). Both carried a rare germline splice site mutation (IVS4+1G>C or c.423+1G>C) in intron 4 of the SDHB gene.4 Analysis of the cDNA from lymphocytes of one of the brothers showed that the last 18 codons of exon 4 were spliced out (Figure 4), predicting a shorter SDHB protein. This mutation was also present in the patients' mother and a sister; both had negative examinations for GIST and paraganglioma.

cDNA studies (RT-PCR) of the PGL146002 (family CTR02) SDHB c.423+1G>C mutation; the predicted protein was shorter but remained in frame.
In the third kindred (B8893/T8231), the proband, now 50 years of age, had a para-aortic paraganglioma and a stomach GIST. At age 26 years, she had severe hypertension and increased urinary content of catecholamines and vanillylmandelic acid. A left nephrectomy and adrenalectomy was performed. The resected specimen included three extra-adrenal paragangliomas. The patient had a novel germline 45_46insCC mutation in exon 1 of the SDHB gene that caused a frameshift and premature termination at codon 76 of the predicted SDHB protein. Allelic marker analysis suggested loss of the SDHB wild-type allele in both the GIST and the paraganglioma from the proband (Figure 5). The proband inherited the mutation from her father who died at the age of 75 years from a poorly differentiated lung adenocarcinoma associated with lung and pleura fibrosis and silicosis: the SDHB mutation was present in the father's normal lung tissue (data not shown).

Loss-of-heterozygosity analysis revealed loss of the SDHB normal allele (the haplotypes segregating with the non-mutant allele were confirmed by analysis of multiple family members) in both a GIST and a para-aortic paraganglioma from patient B8893/T8231.
There were two mutations in the SDHC and one in the SDHD genes. In the fourth family, the proband was a 29-year-old woman (PGL214) who was operated for a pancreatic, non-functional paraganglioma and, a year later, for a large retroperitoneal GIST. A novel, germline nonsense mutation in the SDHC gene (c.43C>T; p. Arg15X) was found.9 The activity of the succinate dehydrogenase-dependent SCCR in the GIST of the proband was significantly decreased with normal COX and QCCR activities;9 there was loss of the wild-type SDHC sequence (data not shown), demonstrating (for the first time in a GIST and, indeed, in any tumor) that ablation of SDHC can cause a severe reduction of succinate dehydrogenase activity.
In the fifth family, the proband (CTRS8), a 20-year-old female, had a gastric GIST and a non-functioning, abdominal, extra-adrenal paraganglioma; she had a rare germline intronic mutation in the SDHC gene (IVS5+1G>A or c.405+1G>A).6 cDNA analysis revealed the transcriptional effect of this splice-site substitution: the mutant allele resulted in a sequence in which exon 5 was spliced out (Figure 6), resulting in a frameshift and a stop codon in the 3′ untranslated region of the gene. More than 75% of the patient's GIST cells showed allelic losses of the SDHC gene locus,9 although the normal sequence was still detectable in the chromatograms (data not shown). The patient inherited the mutation from her apparently clinically healthy 50-year-old mother.

cDNA studies (RT-PCR) of the CTRS8 SDHC IVS5+1G>A mutation: the out-of-frame exon 6 translated directly after exon 4 sequence would predict a stop codon after 36 amino-acids coded by the 3′ region of the SDHC gene.
In the sixth family, the proband (SDH9/04:MB00205) was a 19-year-old male patient who presented with melena from a stomach GIST. At age 21, the patient had a right carotid body tumor and a left adrenal pheochromocytoma; a left glomus jugular tumor was excised a year later. At age 32 he presented with metastatic paraganglioma. Sequencing of germline DNA for the genes encoding succinate dehydrogenase subunits showed a novel germline deletion of a single base at nucleotide 57 of the SDHD gene (c.57delG).9 The mutation resulted in shift in the predicted translation frame of the encoded mRNA from codon 20 to a premature stop codon at position 85. There was no tumor DNA from this patient available; although his lesions were reported to be metastatic, we were not able to prove that genetically.
Because coding sequence mutations were not identified by sequencing the SDHB, SDHC and SDHD genes in the remaining three families, their DNA was also searched for deletions of the respective genes and/or their loci. This analysis revealed a large germline SDHB gene deletion (Figure 7) in another patient who first presented at the age of 9 years with multiple GISTs, and at age 16 had multiple, retroperitoneal paragangliomas. His mother had a paraganglioma but no other information or tumor DNA was available.

(a) SNP intensity in the SDHB region. The value is a normalized ratio in a log2 scale (−0.5 corresponds to loss deletion of one allele). (b) Semi-quantitative duplex PCR of SDHB exons compared to GAPDH exon 8.
Additional LOH studies
The above studies documented SDHB and SDHC allelic losses in all the tumors that had mutations in the respective genes and were available for study (families 1–6), suggesting that these gene defects act in a recessive manner. Four paragangliomas and three GISTs were studied from these families; Supplementary Figure 1 shows the DNA of the tumor showing only the mutant SDHB sequence. Similarly, Figure 5 shows consistent SDHB locus losses in samples (a GIST and a paraganglioma) from the third family. Figure 8a shows loss of the SDHC normal allele in tumor samples from a GIST from the proband of the fifth family. In a recent study, we also showed SDHC mutations and allelic losses that were accompanied by deficient mitochondrial respiratory chain function in family PGL214.9

(a) Loss of the normal allele of the chromosome 1 SDHC locus in a GIST from CTRS8; the mother and the patient, who are both carriers of the SDHC IVS5+1G>A mutation are heterozygotes for the D1S844 a and b alleles, whereas the two tumor samples have retained the b allele. (b) SDHC locus losses among 22 tumors with no SDHC mutations from patients with Carney triad and related conditions: 27% (6/22) of the tumors show at least one allelic marker loss in the vicinity of the SDHC gene.
Finally, we also examined GISTs from other patients collected from our consortium that did not have SDHB and SDHC mutations for losses at the loci of these genes; the frequency of SDHB and SDHC loci losses among 22 tumors (10 GISTs, 10 PGLs and 2 other) ranged between 27 and 40% (Figure 8b). Although we did not investigate the SDHD locus, these data are consistent with those of others for sporadic paragangliomas; GISTs have not been investigated for the respective loci, but both chromosomes 1 and 11 are among those that are frequently reported as altered in genetic investigations by comparative genomic hybridization.18
Discussion
Although rare, GISTs are the most common intramural mesenchymal tumors of the gastrointestinal tract (5000 new cases/year in the United States).18 Median age at diagnosis is around 60 years; the overall 5-year survival is 40% on large tumors with high mitotic index.18, 19, 20 However, GISTs in children or young adults tend to be clinically different and are often part of familial syndromes.18, 20 GISTs originate from stem cells with characteristics of the interstitial cells of Cajal (ICCs), the pacemaker cells which regulate peristalsis in the digestive tract.19 In common with ICCs, up to 95% of GISTs express the receptor tyrosine kinase KIT (CD117) and the hematopoietic progenitor cell antigen marker CD34.19 About 75–80% and 7% of GISTs harbor somatic gain-of-function mutations in the KIT gene10 and in the functionally related PDGFRA gene,20, 21 respectively.
Germline missense mutations or small in-frame deletions of the KIT or PDFGRA genes cause most familial cases of GIST: to date, germline mutations in these genes have been identified in more than 14 families.1, 10, 11, 12 GISTs in these families are characterized by development at multiple primary sites and presentation at a median age of 46 years; polyclonal diffuse hyperplasia of ICCs within the myenteric plexus is considered the primary effect of KIT constitutive activation.10, 22, 23 Other abnormalities may be present in patients with familial GISTs: for example, patients with germline KIT exon 11 mutations often show cutaneous hyperpigmentation in the perineum, face, neck, digits, axillae, groin and knees and less frequently urticaria pigmentosa or diffuse cutaneous mastocytosis in infancy.10, 23 Patients with germline PDGFRA mutations present with multiple gastric GISTs or the condition known as intestinal neurofibromatosis.11, 12
Mutations in KIT and PDGFRA, either somatic or germline, are mutually exclusive events which result in the activation of common signaling pathways including the PI3K/AKT anti-apoptotic, JAK/STAT3 transcriptional and Ras/MAPK mitogenic pathways that respond to STI571 and its analogs.10, 21, 23 The existence of GISTs that lack detectable somatic mutations in either KIT or PDGFRA suggests that a different signaling pathway of tumorigenesis may be involved in the pathogenesis of these neoplasms.
Here we studied the identified germline loss-of-function mutations in the SDHB, SDHC and SDHD genes9 in patients having a recently described familial syndrome known as the dyad of ‘paraganglioma and gastric stromal sarcoma’,2 referred to as the ‘Carney–Stratakis syndrome’.3 The patients presented at a young age (median age 19 years; Table 1). Their tumors displayed losses of the normal allele of the succinate dehydrogenase subunit genes and they (or their relatives) also had paragangliomas, often at multiple sites,2, 7, 9, 17 which prompted us to investigate the sequence of the succinate dehydrogenase subunit genes in germline DNA from these patients.
Three of the mutations we identified had been described before in sporadic or familial pheochromocytomas or extra-adrenal paragangliomas4, 5, 6, 8, 14 (Table 4). A I127S missense mutation of the SDHB gene in a patient with a paraganglioma, who had a family history of paragangliomas, pheochromocytoma and whose maternal grandfather reportedly had a GIST, was also recently described.7 A review of the reported SDHB genetic changes and the ones described here does not reveal an obvious genotype–phenotype correlation (Table 4); however, SDHC mutations are generally rare and have only been found in head and neck paragangliomas.6 Our two patients with SDHC mutations and the dyad (PGL214 and CTRS8) are the first cases with abdominal paragangliomas caused by mutations of this gene.
In one case, we had twins presenting one with a GIST and another with a paraganglioma (family CTR02, extensively described in Baccon-Gibod et al17); although technically for each one of these two patients the diagnosis of the dyad cannot be given, the identification of a common genetic defect supports the probable diagnosis of this syndrome. This family also shows the clinical heterogeneity among our patients: in CTR02, two older patients (the mother and an older sibling) are carriers and unaffected; and from the two affected carriers, one twin had a paraganglioma and the other a GIST.
The absence of KIT or PDGFRA somatic mutations and occurrence of loss of one of the succinate dehydrogenase subunits in GISTs from patients with the dyad, suggests that a deficient mitochondrial tumor suppressor gene pathway is responsible for tumor formation and not constitutively active tyrosine kinases. Succinate dehydrogenase is thus potentially a new molecular target for STI571-unresponsive GISTs. Indeed, gene expression studies in sporadic GISTs without KIT or PDGFRA mutations revealed upregulation of genes linked to the mitochondrial tumor suppressor gene pathway including VEGF,24 a known target of the HIF1 transcription factor.25 As succinate dehydrogenase-dependent tumorigenesis has been associated with an increase in HIF1α nuclear protein,26 it is not unreasonable to speculate that the development of non-KIT and non-PDGFRA-associated GISTs may be associated with abnormal regulation of the hypoxic or pseudo-hypoxic pathway.
In conclusion, familial GISTs may be caused by mutations of the succinate dehydrogenase subunit genes SDHB, SDHC and SDHD, and abdominal paragangliomas associated with GISTs may be caused uniquely by SDHC mutations. Although we do not recommend at this point that all patients with succinate dehydrogenase subunit mutations are screened for GISTs by imaging studies, we do suggest that a careful medical and family history regarding gastrointestinal symptoms and/or tumors should be recorded in every patient with such germline genetic defects. The present study impacts genetic counseling of patients with GISTs or paragangliomas, has implications on the understanding of GIST formation and/or progression, and may lead to new therapies for these tumors.
References
Rubin BP : Gastrointestinal stromal tumours: an update. Histopathology 2006; 48: 83–96.
Carney JA, Stratakis CA : Familial paraganglioma and gastric stromal sarcoma: a new syndrome distinct from the Carney triad. Am J Med Genet 2002; 108: 132–139.
Daum O, Vanecek T, Sima R, Michal M : Gastrointestinal stromal tumor: update. Klinicka Onkologie 2006; 19: 203–211.
Amar L, Bertherat J, Baudin E et al: Genetic testing in pheochromocytoma or functional paraganglioma. J Clin Oncol 2005; 23: 8812–8818.
Brouwers FM, Eisenhofer G, Tao JJ et al: High frequency of SDHB germline mutations in patients with malignant catecholamine-producing paragangliomas: implications for genetic testing. J Clin Endocrinol Metab 2006; 91: 4505–4509.
Schiavi F, Boedeker CC, Bausch B et al: Predictors and prevalance of paraganglioma syndrome associated with mutations of the SDHC gene. JAMA 2005; 294: 2057–2063.
Bolland M, Benn D, Croxson M et al: Gastrointestinal stromal tumour in succinate dehydrogenase subunit B mutation-associated familial phaeochromocytoma/paraganglioma. ANZ J Surg 2006; 76: 763–764.
Neumann HP, Pawlu C, Peczkowska M et al: European-American paraganglioma study group. Distinct clinical features of paraganglioma syndromes associated with SDHB and SDHD gene mutations. JAMA 2004; 292: 943–951.
Passini B, McWhinney SR, Bei T et al: Germline mutations of the genes encoding succinate dehydrogenase subunits (SDHB, SDHC, and SDHD) cause a familial form of gastrointestinal stromal tumors. N Engl J Med (in press).
Hirota S, Isozaki K, Moriyama Y et al: Gain-of-function mutations of c-kit gene in gastrointestinal stromal tumors. Science 1998; 279: 577–580.
Chompret A, Kannengiesser C, Barrois M et al: PDGFRA germline mutation in a family with multiple cases of gastrointestinal stromal tumor. Gastroenterology 2004; 126: 318–321.
de Raedt T, Cools J, Debiec-Rychter M et al: Intestinal neurofibromatosis is a subtype of familial GIST and results from a dominant activating mutation in PDGFRA. Gastroenterology 2006; 131: 1907–1912.
Bertherat J, Groussin L, Sandrini F et al: Molecular and functional analysis of PRKAR1A and its locus (17q22-24) in sporadic adrenocortical tumors: 17q losses, somatic mutations, and protein kinase A expression and activity. Cancer Res 2003; 63: 5308–5319.
Benn DE, Gimenez-Roqueplo AP, Reilly JR et al: Clinical presentation and penetrance of pheochromocytoma/paraganglioma syndromes. J Clin Endocrinol Metab 2006; 91: 827–836.
Bénit P, Goncalves S, Philippe Dassa E, Brière JJ, Martin G, Rustin P : A three-shot assay for the measurement of the five respiratory chain complexes in minute amount of biological samples. Clin Chim Acta 2006; 374: 81–86.
McWhinney SR, Pilarski RT, Forrester SR et al: Large germline deletions of mitochondrial complex II subunits SDHB and SDHD in hereditary paraganglioma 1. J Clin Endocrinol Metab 2004; 89: 5694–5699.
Boccon-Gibod L, Boman F, Boudjema S, Fabre M, Leverger G, Carney AJ : Separate occurrence of extra-adrenal paraganglioma and gastrointestinal stromal tumors in monozygotic twins: probable familial Carney syndrome. Pediatr Dev Pathol 2004; 7: 380–384.
Parfitt JR, Streutker CJ, Riddell RH, Driman DK : Gastrointestinal stromal tumors: a contemporary review. Pathol Res Pract 2006; 202: 837–847.
Kindblom LG, Remotti HE, Aldenborg F, Meis-Kindblom JM : Gastrointestinal pacemaker cell tumor (GIPACT): gastrointestinal stromal tumors show phenotypic characteristics of the interstitial cells of Cajal. Am J Pathol 1998; 152: 1259–1269.
Miettinen M, El-Rifai W, HL Sobin L, Lasota J : Evaluation of malignancy and prognosis of gastrointestinal stromal tumors: a review. Hum Pathol 2002; 33: 478–483.
Heinrich MC, Corless CL, Duensing A et al: PDGFRA activating mutations in gastrointestinal stromal tumors. Science 2003; 299: 708–710.
Kang HJ, Nam SW, Kim H et al: Correlation of KIT and platelet-derived growth factor receptor alpha mutations with gene activation and expression profiles in gastrointestinal stromal tumors. Oncogene 2005; 24: 1066–1074.
Antonescu CR : Gastrointestinal stromal tumor (GIST) pathogenesis, familial GIST, and animal models. Semin Diagn Pathol 2006; 23: 63–69.
Schmahl J, Raymond CS, Soriano P : PDGF signaling specificity is mediated through multiple immediate early genes. Nat Genet 2007; 39: 52–60.
Selak MA, Armour SM, MacKenzie ED et al: Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-alpha prolyl hydroxylase. Cancer Cell 2005; 7: 77–85.
Briere JJ, Favier J, Benit P et al: Mitochondrial succinate is instrumental for HIF1alpha nuclear translocation in SDHA-mutant fibroblasts under normoxic conditions. Hum Mol Genet 2005; 14: 3263–3269.
Acknowledgements
This work was supported by NIH intramural project Z01-HD-000642-04 to Dr CA Stratakis; and, in part, by a Bench-to-Bedside Award from the Office of Rare Disorders (ORD) and the National Institute of Child Health and Human Development (NICHD), NIH, to Drs Stratakis and Eng for the study of the ‘Genetics of inherited paragangliomas and gastric stromal tumors associated with adrenal and other tumors’ and by a grant to Anne-Paule Gimenez-Roqueplo by the GIS-Institut des Maladies Rares for the PGL.NET network. The work of B Pasini and B Ferrando has been supported by Compagnia di San Paolo (Turin) and AIRC (Associazione Italiana Ricerca sul Cancro). Dr C Eng is a recipient of the Doris Duke Distinguished Clinical Scientist Award. The authors are grateful to Dr M Beghini, Dr A Aiello and M Gobbo for technical assistance, and to Prof M Aglietta and Prof M Papotti for reviewing part of the the clinical and pathological data.
Author information
Authors and Affiliations
Corresponding author
Additional information
From the Carney Triad and Carney-Stratakis Dyad Consortium.
Supplementary Information accompanies the paper on European Journal of Human Genetics website (http://www.nature.com/ejhg)
Supplementary information
Rights and permissions
About this article
Cite this article
Pasini, B., McWhinney, S., Bei, T. et al. Clinical and molecular genetics of patients with the Carney–Stratakis syndrome and germline mutations of the genes coding for the succinate dehydrogenase subunits SDHB, SDHC, and SDHD. Eur J Hum Genet 16, 79–88 (2008). https://doi.org/10.1038/sj.ejhg.5201904
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.ejhg.5201904
Keywords
This article is cited by
-
Management of phaeochromocytoma and paraganglioma in patients with germline SDHB pathogenic variants: an international expert Consensus statement
Nature Reviews Endocrinology (2024)
-
TKI Treatment Sequencing in Advanced Gastrointestinal Stromal Tumors
Drugs (2023)
-
Observational study of population genomic screening for variants associated with endocrine tumor syndromes in a large, healthcare-based cohort
BMC Medicine (2022)
-
The Management of Unresectable, Advanced Gastrointestinal Stromal Tumours
Targeted Oncology (2022)
-
Abdominelle Weichgewebstumoren
Die Pathologie (2022)

{kind=link}