
Abstract
The charisma of top vertebrate predators is often used by conservationists as a lever for financial support1,2, to raise environmental awareness2,3 and in planning protected areas4,5,6 — a strategy that has been criticized3,5,7. Here we use information collected from five raptor species that differ widely in their diet and habitat associations to show that sites occupied by these predators are consistently associated with high biodiversity. The biodiversity at these sites is more extensive than it is at sites selected at random, or at sites occupied by species from lower down the trophic pyramid (insectivorous or herbivorous species, for example). Our results indicate that conservation focusing on top predators can be ecologically justified because it delivers broader biodiversity benefits.
Similar content being viewed by others

Main
We surveyed five species of diurnal and nocturnal raptors in the Italian Alps during the breeding season: the goshawk (Accipiter gentilis), pygmy owl (Glaucidium passerinum), Tengmalm's owl (Aegolius funereus), tawny owl (Strix aluco) and scops owl (Otus scops) (for site locations, see supplementary information). The diversity of birds, trees and butterflies was used as a surrogate measure of biodiversity. For each raptor species, we compared the biodiversity recorded at 25 sites occupied for breeding with that at three other types of site: 25 sites randomly chosen in comparable habitat (spatial-control sites); 25 breeding sites of a randomly selected species from a lower trophic level (taxonomic-control sites A); and 25 breeding sites of a species from a lower trophic level that has specialized ecological requirements (taxonomic-control sites B). (For methods, see supplementary information.)
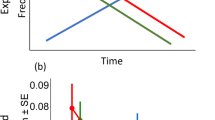
Compared with the three types of control site, locations occupied by top predators had greater numbers and more diversity of avian species, vulnerable avian species, butterfly species and tree species (for all raptor species and for all comparisons, t⩾3.14, P<0.05, except for one comparison, for which t=2.96, P<0.07; Fig. 1a–c). By contrast, none of the biodiversity estimates collected at sites occupied by species from lower trophic levels was higher than those collected at their associated spatial-control sites (for all species, t⩽2.45, P⩾0.16; Fig. 1a–c).

Red bars, breeding sites; blue bars, randomly selected spatial-control sites. Values represent averages ±1 s.e. a, Numbers of all avian species. b, Numbers of avian species classified as vulnerable. c, Numbers of tree species. d, Percentage of maximum attainable avian-species richness in a hypothetical system of protected areas, as estimated by gap analysis7,8.
We also did a simulation exercise (gap analysis7,8; see supplementary information) that selects a network of protected areas by maximizing the overall level of biodiversity representation while minimizing the number of protected areas included in the network. This showed that the number of sites required to include all avian species, or all vulnerable avian species, in a hypothetical protected system was lower when using sites occupied by top predators than when using any type of control site (t⩾2.89, P⩽0.05). The difference between sites occupied by species from lower trophic levels and their associated spatial control sites was not significant (t⩽1.64, P⩾0.14).
Furthermore, the efficiency of each protected-area system (expressed as the percentage of maximum attainable biodiversity) was higher for top-predator sites than for any type of control site (t⩾3.97, P<0.01; Fig. 1d), whereas it did not differ between sites that were occupied by lower-trophic-level species and their associated spatial-control sites (t⩽1.36, P⩾0.21; Fig. 1d). On average, networks planned using lower-trophic-level species represented 72% of the maximum species richness and 76% of the maximum richness of vulnerable species in each sample, compared with 94% and 93% for networks based on the presence of top predators.
Our results are evidence of a tight association, at least in some biological systems, between apex predators and high biodiversity, which may justify the strategic economic exploitation of top-predator species on ecological grounds. More information from other systems will be needed before the powerful top predators can be dismissed from the conservation arena as unscientific tools.
References
White, P. C. L., Gregory, K. W., Lindley, P. J. & Richards, G. Biol. Conserv. 82, 345–354 (1997).
Gittleman, J. L., Funk, S. M., Macdonald, D. & Wayne, R. K. (eds) Carnivore Conservation (Cambridge Univ. Press, Cambridge, 2001).
Entwistle, A. & Dunstone, N. (eds) Priorities for the Conservation of Mammalian Diversity: Has the Panda Had its Day? (Cambridge Univ. Press, Cambridge, 2000).
Murphy, D. D. & Noon, B. R. Ecol. Appl. 2, 3–17 (1992).
Andelman, S. J. & Fagan, W. F. Proc. Natl Acad. Sci. USA 97, 5954–5959 (2000).
Carroll, C., Noss, R. F. & Paquet, P. C. Ecol. Appl. 11, 961–980 (2001).
Kerr, J. T. Conserv. Biol. 11, 1094–1100 (1997).
Jeggins, M. D. Landscape Ecol. 15, 5–20 (2000).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
About this article
Cite this article
Sergio, F., Newton, I. & Marchesi, L. Top predators and biodiversity. Nature 436, 192 (2005). https://doi.org/10.1038/436192a
Published:
Issue Date:
DOI: https://doi.org/10.1038/436192a
This article is cited by
-
Wintering raptor species distribution in a semiarid Mediterranean region: the relevance of lowlands and open habitats as stopover sites
European Journal of Wildlife Research (2024)
-
Discovery of a colossal slickhead (Alepocephaliformes: Alepocephalidae): an active-swimming top predator in the deep waters of Suruga Bay, Japan
Scientific Reports (2021)
-
Raptor breeding sites indicate high plant biodiversity in urban ecosystems
Scientific Reports (2021)
-
Habitat loss causes long extinction transients in small trophic chains
Theoretical Ecology (2021)
-
Platypus predation has differential effects on aquatic invertebrates in contrasting stream and lake ecosystems
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
