
Similar content being viewed by others

Liu et al. reply
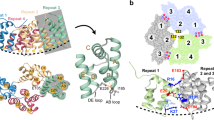
We have shown that soluble TALL-1 (sTALL-1; residues 134–285) with a Flag tag, secreted from 293 cell lines, spontaneously forms a virus-like cluster in the culture medium under physiological conditions1. Also, Flag-tagged proteins dissociate into trimers and monomers at low pH, ruling out the possibility of random aggregation that is suggested by Zhukovsky and colleagues.
The pH dependence of the sTALL-1 structure is consistent with the presence of a histidine residue in the flap region1, the removal of which prevents the oligomerization of sTALL-1 and abolishes the activity of the ligand in transfection and in B-cell-stimulation assays1. However, this truncated version of the molecule still binds to its receptor1, indicating that the missing flap region is not involved in receptor binding2. The structure of the truncated molecule is the same as that of native sTALL-1, apart from the missing flap region2.
It is true that some His6-tagged proteins are susceptible to aggregation caused by divalent metals. We tested whether this could be the case for our His6-tagged sTALL-1 protein by adding 100 mM EDTA, pH 7.5, at the different stages of purification. We found that it did not affect oligomerization. Note also that the His6 tag is far away from the trimer–trimer association region in our structure1.
To prevent any perturbation of the oligomerization properties of sTALL-1 by the His6 or Flag tags, we overexpressed an untagged version of sTALL-1 (residues 134–285) in Escherichia coli. This sTALL-1 (prepared and characterized with the help of Zhongzhou Chen) was purified on a Q-Sepharose column in buffer (20 mM Tris-HCl) at pH 7.2, concentrated in an Amicon Ultra unit (Millipore) and applied to Superdex-200 (150 mM NaCl, 20 mM Tris-HCl) at pH 8.0. sTALL-1 emerged as a multimer in the void volume (data not shown). The buffer containing this sample was then adjusted to pH 6.0, equilibrated for 2 hours and reapplied to the Superdex-200 column in the same buffer at pH 6.0.
The elution profile consisted of a single sharp peak: the molecular size of the material in this fraction was estimated from the elution positions of three Mr standards (blue dextran, 2,000,000 (2,000K); ferritin, 440K; and ovalbumin, 43K) under the same conditions. The pure sTALL-1 protein eluted at a position corresponding to a trimer. Amino-terminal sequencing analysis confirmed that this was the correct protein (data not shown).
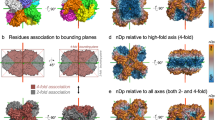
Other crystal structures3,4 reported for trimeric sTALL-1 were obtained with proteins that had been crystallized at low pH (pH 4.5 and 6.0, respectively), conditions under which the trimer should be the only species present1. Zhukovsky et al. used native sTALL-1, with and without tags, in their assays. As these should both be transformed to 60-mers under physiological conditions, it is not surprising that these forms are active. If, by contrast, they had used a flapless version of the protein, which does not oligomerize, we predict that it would have been inactive. We contest that neither the His6 tag nor the Flag tag promotes or blocks the formation of 60-mers by sTALL-1 and that it is the 60-mer, rather than a trimer or some other oligomer, that is the functional unit.
Other data indicate that further oligo-merization of ligand trimers (multiple valences) or mulitple trimers are required to activate the signal-transduction pathway of the TNF superfamily. For example, TNF receptors can oligomerize on the cell surface without bound ligand5. TNF-receptor-associated factors (TRAF), which mediate downstream signal transduction by the TNF family, exist as trimers before TNF ligands are recruited to its receptors6. Ectodysplasin ligand A1 (EDA-A1) and A2 (EDA-A2) require a collagen-like region for their proper function, which suggests that two dimers or higher-order clusters are needed for the activation of these ligands7. Combining with our results with sTALL-1, it is reasonable to suggest that one trimer of the ligand is not enough to trigger signal transduction for TNF family members. Multiple trimers (including trimers in different oligomerization states of increased concentration and valence and hence increased avidity) or a local accumulation of trimers (increasing in concentration alone) on the cell surface may be required for the optimal recruitment of a cluster of receptors to activate signal transduction. This is similar to the supermolecular activation cluster phenomenon of T-cell activation that occurs after binding to antigen-presenting cells8.
References
Liu, Y. et al. Cell 108, 383–394 (2002).
Liu, Y. et al. Nature 423, 49–56 (2003).
Karpusas, M. et al. J. Mol. Biol. 315, 1145–1154 (2002).
Oren, D. A. et al. Nature Struct. Biol. 9, 288–292 (2002).
Chan, F. K. et al. Science 288, 2351–2354 (2000).
Park, Y. C. et al. Nature 398, 533–538 (1999).
Srivastava, A. K. et al. Proc. Natl Acad. Sci. USA 94, 13069–13074 (1997).
Monks, C. R., Freiburg, B. A., Kupfer, H., Sciaty, N. & Kupfer, A. Nature 395, 82–86 (1998).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hong, X., Kappler, J., Liu, Y. et al. Is Tall-1 a trimer or a virus-like cluster?. Nature 427, 414 (2004). https://doi.org/10.1038/427414a
Issue Date:
DOI: https://doi.org/10.1038/427414a
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
