
Silk fibres produced by artificial reeling are superior to those that are spun naturally.
Abstract
Commercial silkworm silk is presumed to be much weaker and less extensible than spider dragline silk, which has been hailed as a 'super-fibre'1,2,3,4. But we show here that the mechanical properties of silkworm silks can approach those of spider dragline silk when reeled under controlled conditions. We suggest that silkworms might be able to produce threads that compare well with spider silk by changing their spinning habits, rather than by having their silk genes altered.
Similar content being viewed by others

Main
Typical commercial silkworm silk from Bombyx mori cocoons has a tensile strength of about 0.5 gigapascals (GPa), a breaking elongation of 15%, and a breaking energy (toughness) of 6 × 104 J kg−1 (ref. 5). Nephila spider dragline silk, on the other hand, typically has a strength of 1.3 GPa, a breaking elongation of 40%, and a toughness of 16 × 104 J kg−1 (ref. 4).
These mechanical measures vary considerably in both types of silk. For spider silk, this variability is due to spinning conditions, which are affected by the spider's body temperature and the speed of drawing4. Silkworm silk is traditionally obtained from a natural cocoon that is spun by the moving silkworm, which accelerates and decelerates its head in arcs that are attached at points that correspond to each change of direction6.
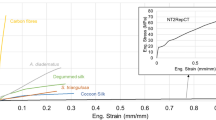
We find that artificial reeling of silk from immobilized silkworms under steady and controlled conditions produces fibres that are superior to naturally spun ones. The silkworm, like the spider4, produces stronger, more brittle fibres at faster spinning speeds, whereas slower speeds lead to weaker, more extensible fibres (Fig. 1).

Stress–strain curves of washed and degummed single-filament silkworm silk (motor-reeled at 25 °C at the indicated speeds), Nephila spider dragline silk (20 mm s−1 at 25 °C) and standard, degummed commercial silk from a cocoon spun by the animal in the natural 'figure of eight'9 at speeds oscillating between 4 and 15 mm s−1 at 20 °C. The area under the stress–strain curve represents the energy that a fibre can take up before breaking, and thus indicates its toughness. Scale bar, 10 µm. Immobilized silkworms (n = 4) were forcibly silked4, each providing 3–6 single filaments, which were tested in a stretching rig (force resolution, 30 µN; time resolution, <5 ms for 1 mN; strain rate, 50% per min)4; further details are available from the authors. For silk 'degumming', a traditional aqueous solution standard wash of 1% sodium hydrogen carbonate10 was used, which led to a 30–40% reduction in fibre diameter. Inset, unwashed native silkworm silk (photo courtesy of Giuliano Freddi).
Force-drawn silkworm fibres compared favourably with Nephila spider dragline silk. For example, slow spinning at 4 mm s−1 gave a breaking elongation for Bombyx silk that is comparable to that of spider silk (37% and 35%, respectively). Faster spinning (13 mm s−1) gave a breaking energy approaching that of Nephila silk (12 × 104 J kg−1 for Bombyx; versus 16 × 104 J kg−1 for Nephila), although the breaking strength remained lower (0.7 GPa compared with 1.3 GPa). At even greater spinning speeds, silk toughness decreased, mainly because of loss of extensibility (Fig. 1).
Other differences between silkworm and spider silk may be linked to specific differences in the composition of the principal silk molecules2, as well as to their arrangement during and after spinning1. Depending on reeling speed, Bombyx silk is either strong or stretchable (Fig. 1), whereas spider dragline silk typically combines the two at the speeds that are characteristic of web construction4 (10–20 mm s−1).
Although spider silk fibres are studied in their native state4, silkworm silk must first be washed7 to remove most of the sericin gum coating the fibroin filaments from the 'bave' thread (Fig. 1, inset). Washing extraction is necessary for the thread to be unravelled from the cocoon (Fig. 2), and is generally assumed to weaken the thread8. Our experiments (each run four times) on silkworm silk filaments reeled under controlled conditions (12–24 filaments per treatment) showed that spinning conditions affect the silk's material properties more than washing.

Changing the spinning conditions can improve the silk's quality.
For example, washing experimental fibres reeled at 4 mm s−1 changed neither the force required to break a thread (washed, 36 ± 2 mN; native, 38 ± 4 mN, P = 0.34, t-test) nor its breaking elongation (washed, 37 ± 3%; native, 39 ± 4%, P = 0.53). However, 'degumming' significantly increased viscoelastic recovery (washed, 40 ± 3%; native, 29 ± 2%, P < 0.001), as shown by the loading–unloading cycles of single fibres stretched to 50% of typical breaking elongation values. Thus, more than anything else, washing increases the elasticity of fibroin filaments, presumably because of the combined action of sericin removal and water plasticization.
Our findings indicate that the mechanical properties of Bombyx silk, like those of spider silk4, depend crucially on spinning conditions — silkworm silks can be made stronger, stiffer and more extensible simply by adjusting the harvesting parameters. If we could reel silk straight from the silkworm, as from spiders, or if larvae could be bred to spin their cocoons faster and more evenly (with pretreatment washing appropriately modified), then the silkworm would produce fibres that might well give natural and artificially spun spider silks — genetically modified or not — a good run for their money.
References
Vollrath, F. & Knight, D. P. Nature 410, 541–548 (2001).
Kaplan, D. L., Adams, W. W., Viney, C. & Farmer, B. L. Silk Polymers: Materials Science and Biotechnology (ACS, Washington, 1994).
Lazaris, A. et al. Science 295, 472–476 (2002).
Vollrath, F., Madsen, B. & Shao, Z. Proc. R. Soc. Lond. B 268, 2339–2346 (2001).
Wilding, M. A. & Hearle, J. W. S. in Polymeric Materials Encyclopaedia Vol. 11 (ed. Salamone, J. C.) 8307–8322 (CRC, Boca Raton, Florida, 1996).
Miura, M., Pan, Z. J., Aoyama, S., Morikawa, H. & Mochizuki, S. J. Seric. Sci. Jpn 67, 51–56 (1998).
Perez-Rigueiro, J., Elices, M. J., Llorca, J. & Viney, C. J. Appl. Polymer Sci. 82, 1928–1935 (2001).
Perez-Rigueiro, J., Viney, C., Llorca, J. & Elices, M. Polymer 41, 8433–8439 (2000).
Wiedbrauck, J. Z. Tierpsychol. 12, 176–202 (1955).
Monti, P., Freddi, G., Bertoluzza, A., Kasai, N. & Tsukada, M. J. Raman Spectrosc. 29, 297–304 (1998).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Shao, Z., Vollrath, F. Surprising strength of silkworm silk. Nature 418, 741 (2002). https://doi.org/10.1038/418741a
Issue Date:
DOI: https://doi.org/10.1038/418741a
This article is cited by
-
Replicating shear-mediated self-assembly of spider silk through microfluidics
Nature Communications (2024)
-
Hierarchically structured bioinspired nanocomposites
Nature Materials (2023)
-
Strong yet flexible ceramic aerogel
Nature Communications (2023)
-
Photonic structures in radiative cooling
Light: Science & Applications (2023)
-
Bioinspired Hard–Soft Interface Management for Superior Performance in Carbon Fibre Composites
Fibers and Polymers (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
