
Abstract
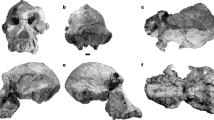
Similarities of the skull are commonly used to support hypotheses of ancestor–descendant relationships between fossil and living ape genera, especially between the late Miocene apes Sivapithecus and Dryopithecus from Eurasia and the living orang-utan (Pongo) from Borneo and Sumatra1,2,3,4. Yet determining whether craniofacial traits shared by extant and Miocene apes are primitive or derived is severely hampered by the rarity of well-preserved fossil crania, particularly of early members of their closest outgroup, the Old World monkeys (Cercopithecoidea). The discovery of a complete and undistorted skull of Victoriapithecus at middle Miocene deposits from Maboko Island, Kenya, provides evidence of intact cranial-vault and basicranial morphology, brain size and craniofacial hafting for a primate from between 32 and 7 million years ago. Victoriapithecus represents a branch of Old World monkey that is intermediate between extant cercopithecids (Colobinae and Cercopithecinae) and the common ancestor they shared with apes (Hominoidea)5,6,7,8. The skull preserves traits widely thought to be derived for extant and fossil members of a proposed Sivapithecus/Pongo clade, but which now appear to be primitive features of ancestral Old World higher primates in general.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

References
Pilbeam, D. R. New hominoid skull material from the Miocene of Pakistan. Nature 295, 232–234 (1982).
Brown, B. & Ward, S. in Orang-utan Biology(ed. Schwartz, J. H.) 247–260 (Oxford Univ. Press, New York, (1988)).
Moya-Sola, S. & Kohler, M. Recent discoveries of Dryopithecus shed new light on evolution of great apes. Nature 365, 543–545 (1993).
Moya-Sola, S. & Kohler, M. New partial cranium of Dryopithecus Lartet, 1863 (Hominoidea, Primates) from the upper Miocene of Can Llobateres, Barcelona, Spain. J. Hum. Evol. 29, 101–139 ((1995)).
Von Koenigswald, G. H. R. Miocene Cercopithecoidea and Oreopithecoidea from the Miocene of East Africa. Foss. Vert. Afr. 1, 39–51 (1969).
Benefit, B. R. & McCrossin, M. L. Ancestral facial morphology of Old World higher primates. Proc. Natl Acad. Sci. USA 88, 5267–5271 (1991).
Benefit, B. R. The permanent dentition and phylogenetic position of Victoriapithecus from Maboko Island, Kenya. J. Hum. Evol. 25, 83–172 (1993).
Benefit, B. R. Phylogenetic, paleodemographic and taphonomic implications of Victoriapithecus deciduous teeth from Maboko, Kenya. Am. J. Phys. Antropol. 95, 277–331 (1994).
Feibel, C. S. & Brown, F. H. Age of the primate-bearing deposits on Maboko Island, Kenya. J. Hum. Evol. 21, 221–225 (1991).
Andrews, P., Meyer, G., Pilbeam, D. R., Van Couvering, J. A. & Van Couvering, J. A. H. The Miocene fossil beds of Maboko Island, Kenya: Geology, age, taphonomy and paleontology. J. Hum. Evol. 10, 35–48 (1981).
Stromer, E. Mitteilungen uber die Wirbeltierreste aus dem Mittelpliocan des Natrontales (Agypten). 1. Affen. Z. Deutsch. Geol. Gesell. Abh. 65, 349–361 (1913).
Harrison, T. New postcranial remains of Victoriapithecus from the middle Miocene of Kenya. J. Hum. Evol. 18, 3–54 (1989).
Radinsky, L. The fossil evidence of anthropoid brain evolution. Am. J. Phys. Anthropol. 41, 15–28 (1974).
Martin, R. D. Primate Origins and Evolution(Princeton Univ. Press, Princeton, NJ, (1990)).
Benefit, B. R. & McCrossin, M. L. The facial anatomy of Victoriapithecus and its relevance to the ancestral cranial morphology of Old World monkeys and apes. Am. J. Phys. Anthropol. 92, 329–370 (1993).
Vogel, C. Morphologische studien am gesichtschadel Catarrhiner primaten. Biblio Primatol. 4, 1–226 (1966).
Harrison, T. The phylogenetic relationships of the early catarrhine primates: A review of the current evidence. J. Hum. Evol. 16, 41–80 (1987).
Simons, E. L. The earliest apes. Sci. Am. 217, 28–35 (1967).
Simons, E. L. New faces of Aegyptopithecus from the Oligocene of Egypt. J. Hum. Evol. 16, 273–289 (1987).
Leakey, R. E. & Leakey, M. G. Anew Miocene hominoid from Kenya. Nature 342, 143–146 (1986).
Leakey, M. G., Leakey, R. E., Richtsmeier, J. T., Simons, E. L. & Walker, A. C. Similarities in Aegyptopithecus and Afropithecus facial morphology. Folia Primatol. 56, 65–85 (1991).
Delson, E. & Andrews, P. in Phylogeny of the Primates: A Multidisciplinary Approach(eds Luckett, W. P. & Szalay, F. S.) 405–446 (Plenum, New York, (1975)).
Strasser, E. & Delson, E. Cladistic analysis of cercopithecid relationships. J. Hum. Evol. 16, 81–99 (1987).
Acknowledgements
Excavations at Maboko were conducted with permission of the Office of the President, Republic of Kenya and in collaboration with the National Museums of Kenya. We thank the field crew (especially B. Onyango, V. Oluoch and S. Gitau) and M. G. Leakey for assistance, and E. Delson, M. Kohler, S. Moya-Sola and D. Pilbeam for comments and advice. This work was supported by NSF, L.S.B. Leakey Foundation, National Geographic Society, Wenner Gren Foundation for Anthropological Research, Fulbright, and Boise Fund.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Benefit, B., McCrossin, M. Earliest known Old World monkey skull. Nature 388, 368–371 (1997). https://doi.org/10.1038/41078
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/41078
This article is cited by
-
Dental Eruption Sequences in Fossil Colobines and the Evolution of Primate Life Histories
International Journal of Primatology (2007)
-
A molecular timescale for vertebrate evolution
Nature (1998)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
