
Abstract
Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) has long been used as a traditional Chinese medicine for dysfunctions of the endocrine system and inflammation conditions. However, the effect of adlay seed on the endocrine system has not yet been reported. In the present study, the effects and the mechanisms of methanolic extract of adlay bran (ABM) on progesterone synthesis in rat granulosa cell were studied. ABM was further partitioned with different solvents including water, 1-butanol, ethyl acetate and n-hexane. Four subfractions named ABM-Wa (water fraction), ABM-Bu (1-butanol fraction), ABM-EA (ethyl acetate fraction) and ABM-Hex (n-hexane fraction) were obtained. ABM-Bu was further fractionated using Diaion HP-20 resin column chromatography with gradient elution. Granulosa cells were prepared from pregnant mare serum gonadotropin-primed immature female rats and challenged with different reagents including human chorionic gonadotropin (hCG 0.5 IU/ml), forskolin (10 μ M), 8-bromo-adenosine-3′,5′-cyclic monophosphate (8-Br-cAMP, 1 mM), A23187 (10 μ M), phorbol 12-myristate 13-acetate (PMA, 0.01 μ M), 25-OH-cholesterol (0.1–10 μ M) and pregnenolone (0.1–10 μ M) in the presence or absence of ABM-Bu (100 μg/ml). The functions of steroidogenic enzyme including protein expression of the steroidogenic acute regulatory protein (StAR) and cytochrome P450 side-chain cleavage enzyme (P450scc) protein were investigated. Expressions of both P450scc and StAR mRNA have also been explored. We found that ABM decreased progesterone production via an inhibition on (1) the cAMP-PKA and PKC signal transduction pathway, (2) P450scc and 3β-hydroxysteroid dehydrogenase (3β-HSD) enzyme activity, (3) P450scc and StAR protein and mRNA expressions and (4) the phosphorylation of ERK1/2 in rat granulosa cells.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 8 print issues and online access
$259.00 per year
only $32.38 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout












Similar content being viewed by others
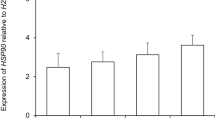
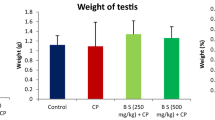
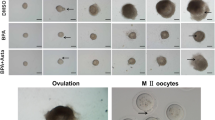
References
Li SZ . Pen's Sao Kang Mu (Compendium of Materia Medica, Systematic Pharmacacopoeia). China, 1596. Wangchao Information Technology Co., Ltd: Huanggang, 2004.
Hidaka Y, Kaneda T, Amino N, Miyai K . Chinese medicine, Coix seeds increase peripheral cytotoxic T and NK cells. Biotherapy 1992; 5 (3): 201–203.
Numata M, Yamamoto A, Moribayashi A, Yamada H . Antitumor components isolated from the Chinese herbal medicine Coix lachryma-jobi. Planta Med 1994; 60 (4): 356–359.
Hung WC, Chang HC . Methanolic extract of adlay seed suppresses COX-2 expression of human lung cancer cells via inhibition of gene transcription. J Agric Food Chem 2003; 51 (25): 7333–7337.
Chang HC, Huang YC, Hung WC . Antiproliferative and chemopreventive effects of adlay seed on lung cancer in vitro and in vivo. J Agric Food Chem 2003; 51 (12): 3656–3660.
Shih CK, Chiang W, Kuo ML . Effects of adlay on azoxymethane-induced colon carcinogenesis in rats. Food Chem Toxicol 2004; 42 (8): 1339–1347.
Kuo CC, Chiang W, Liu GP, Chien YL, Chang JY, Lee CK et al. 2,2′-Diphenyl-1-picrylhydrazyl radical-scavenging active components from adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) hulls. J Agric Food Chem 2002; 50 (21): 5850–5855.
Kuo CC, Shih MC, Kuo YH, Chiang W . Antagonism of free-radical-induced damage of adlay seed and its antiproliferative effect in human histolytic lymphoma U937 monocytic cells. J Agric Food Chem 2001; 49 (3): 1564–1570.
Hsu HY, Lin BF, Lin JY, Kuo CC, Chiang W . Suppression of allergic reactions by dehulled adlay in association with the balance of TH1/TH2 cell responses. J Agric Food Chem 2003; 51 (13): 3763–3769.
Kondo Y, Nakajima K, Nozoe S, Suzuki S . Isolation of ovulatory-active substances from crops of Job's tears (Coix lacryma-jobi L. var. ma-yuen STAPF.). Chem Pharm Bull (Tokyo) 1988; 36 (8): 3147–3152.
McGuffin M, Hornsby C, Upon R, Goldberg A . Botanical Safety Handbook, 1st edn. American Herbal Products Association, CRC-Press: Boca Raton, FL, 1997.
Lauber ME, Picton HM, Begeot M, Momoi K, Waterman MR, Simpson ER . Regulation of CYP11Agene expression in bovine ovarian granulosa cells in primary culture by cAMP and phorbol esters is conferred by a common cis-acting element. Mol Cell Endocrinol 1993; 94 (2): 235–242.
Stocco DM . Intramitochondrial cholesterol transfer. Biochim Biophys Acta 2000; 1486 (1): 184–197.
Stocco CO, Chedrese J, Deis RP . Luteal expression of cytochrome P450 side-chain cleavage, steroidogenic acute regulatory protein, 3beta-hydroxysteroid dehydrogenase, and 20alpha-hydroxysteroid dehydrogenase genes in late pregnant rats: effect of luteinizing hormone and RU486. Biol Reprod 2001; 65 (4): 1114–1119.
Stocco DM . StAR protein and the regulation of steroid hormone biosynthesis. Annu Rev Physiol 2001; 63: 193–213.
Stocco DM . An update on the mechanism of action of the Steroidogenic Acute Regulatory (StAR) protein. Exp Clin Endocrinol Diabetes 1999; 107 (4): 229–235.
Too CK, Weiss TJ, Bryant-Greenwood GD . Relaxin stimulates plasminogen activator secretion by rat granulosa cells in vitro. Endocrinology 1982; 111 (4): 1424–1426.
Hu MC, Guo IC, Lin JH, Chung BC . Regulated expression of cytochrome P-450scc (cholesterol-side-chain clevage enzyme) in cultured cell lines detected by antibody against bacterially expressed human protein. Biochem J 1991; 274: 813–817.
Lin T, Hu J, Wang D, Stocco DM . Interferon-gamma inhibits the steroidogenic acute regulatory protein messenger ribonucleic acid expression and protein levels in primary cultures of rat Leydig cells. Endocrinology 1998; 139 (5): 2217–2222.
Hwang JJ, Lin SW, Teng CH, Ke FC, Lee MT . Relaxin modulates the ovulatory process and increases secretion of different gelatinases from granulosa and theca-interstitial cells in rats. Biol Reprod 1996; 55 (6): 1276–1283.
Tsai SC, Lu CC, Chen JJ, Chiao YC, Wang SW, Hwang JJ et al. Inhibition of salmon calcitonin on secretion of progesterone and GnRH-stimulated pituitary luteinizing hormone. Am J Physiol 1999; 277 (1 Part 1): E49–E55.
Lu SS, Lau CP, Tung YF, Huang SW, Chen YH, Shih HC et al. Lactate stimulates progesterone secretion via an increase in cAMP production in exercised female rats. Am J Physiol 1996; 271 (5 Part 1): E910–E915.
Chen TS, Doong ML, Wang SW, Tsai SC, Lu CC, Shih HC et al. Gastric emptying and gastrointestinal transit during lactation in rats. Am J Physiol 1997; 272 (3 Part 1): G626–G631.
Chen JJ, Chien EJ, Wang PS . Progesterone attenuates the inhibitory effects of cardiotonic digitalis on pregnenolone production in rat luteal cells. J Cell Biochem 2002; 86 (1): 107–117.
Chen JJ, Wang PS, Chien EJ, Wang SW . Direct inhibitory effect of digitalis on progesterone release from rat granulosa cells. Br J Pharmacol 2001; 132 (8): 1761–1768.
Chen JJ, Wang SW, Chien EJ, Wang PS . Direct effect of propylthiouracil on progesterone release in rat granulosa cells. Br J Pharmacol 2003; 139 (8): 1564–1570.
Bradford MM . A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976; 72: 248–254.
Kau MM, Lo MJ, Wang SW, Tsai SC, Chen JJ, Chiao YC et al. Inhibition of aldosterone production by testosterone in male rats. Metabolism 1999; 48 (9): 1108–1114.
Chiao YC, Cho WL, Wang PS . Inhibition of testosterone production by propylthiouracil in rat Leydig cells. Biol Reprod 2002; 67 (2): 416–422.
Ronen-Fuhrmann T, Timberg R, King SR, Hales KH, Hales DB, Stocco DM et al. Spatio-temporal expression patterns of steroidogenic acute regulatory protein (StAR) during follicular development in the rat ovary. Endocrinology 1998; 139 (1): 303–315.
Orly J, Stocco DM . The role of the steroidogenic acute regulatory (StAR) protein in female reproductive tissues. Horm Metab Res 1999; 31 (7): 389–398.
Steel RGD, Torrie JH . Principles and Procedures of Statistic. McGraw-Hill: New York, 1960.
Sokka TA, Hamalainen TM, Kaipia A, Warren DW, Huhtaniemi IT . Development of luteinizing hormone action in the perinatal rat ovary. Biol Reprod 1996; 55 (3): 663–670.
Lauber ME, Kagawa N, Waterman MR, Simpson ER . cAMP-dependent and tissue-specific expression of genes encoding steroidogenic enzymes in bovine luteal and granulosa cells in primary culture. Mol Cell Endocrinol 1993; 93 (2): 227–233.
Danisova A, Scsukova S, Matulova L, Orlicky J, Kolena J . Role of calcium in luteinization stimulator-enhanced progesterone production of porcine granulosa cells. Physiol Res 1995; 44 (3): 185–192.
Tsang BK, Carnegie JA . Calcium-dependent regulation of progesterone production by isolated rat granulosa cells: effects of the calcium ionophore A23187, prostaglandin E2, dl-isoproterenol and cholera toxin. Biol Reprod 1984; 30 (4): 787–794.
Flores JA, Garmey JC, Nestler JE, Veldhuis JD . Sites of inhibition of steroidogenesis by activation of protein kinase-C in swine ovarian (granulosa) cells. Endocrinology 1993; 132 (5): 1983–1990.
He H, Herington AC, Roupas P . Effect of protein kinase C modulation on gonadotrophin-induced granulosa cell steroidogenesis. Reprod Fertil Dev 1995; 7 (1): 83–95.
Johnson AL, Solovieva EV, Bridgham JT . Relationship between steroidogenic acute regulatory protein expression and progesterone production in hen granulosa cells during follicle development. Biol Reprod 2002; 67 (4): 1313–1320.
Hadley ME . Hormone and female reproduction physiology. In: Hadlay ME (ed). Endocrinology, 5th edn. Prentice-Hall International Inc.: Englewood Cliffs, NJ, 2000, pp 445–472.
Clark BJ, Wells J, King SR, Stocco DM . The purification, cloning, and expression of a novel luteinizing hormone-induced mitochondrial protein in MA-10 mouse Leydig tumor cells. Characterization of the steroidogenic acute regulatory protein (StAR). J Biol Chem 1994; 269 (45): 28314–28322.
Clark BJ, Combs R, Hales KH, Hales DB, Stocco DM . Inhibition of transcription affects synthesis of steroidogenic acute regulatory protein and steroidogenesis in MA-10 mouse Leydig tumor cells. Endocrinology 1997; 138 (11): 4893–4901.
Stocco DM, Clark BJ . Role of the steroidogenic acute regulatory protein (StAR) in steroidogenesis. Biochem Pharmacol 1996; 51 (3): 197–205.
Stocco DM . A review of the characteristics of the protein required for the acute regulation of steroid hormone biosynthesis: the case for the steroidogenic acute regulatory (StAR) protein. Proc Soc Exp Biol Med 1998; 217 (2): 123–129.
Moore RK, Otsuka F, Shimasaki S . Role of ERK1/2 in the differential synthesis of progesterone and estradiol by granulosa cells. Biochem Biophys Res Commun 2001; 289 (4): 796–800.
Dewi DA, Abayasekara DR, Wheeler-Jones CP . Requirement for ERK1/2 activation in the regulation of progesterone production in human granulosa-lutein cells is stimulus specific. Endocrinology 2002; 143 (3): 877–888.
Seger R, Hanoch T, Rosenberg R, Dantes A, Merz WE, Strauss III JF et al. The ERK signaling cascade inhibits gonadotropin-stimulated steroidogenesis. J Biol Chem 2001; 276 (17): 13957–13964.
Gyles SL, Burns CJ, Whitehouse BJ, Sugden D, Marsh PJ, Persaud SJ et al. ERKs regulate cyclic AMP-induced steroid synthesis through transcription of the steroidogenic acute regulatory (StAR) gene. J Biol Chem 2001; 276 (37): 34888–34895.
Wang SC . Quantification and comparison of physiological components in adlay seeds. M.S. Thesis, Graduate Institute of Food Science and Technology National Taiwan University, Taipei, Taiwan, ROC 2002.
Ohno S, Shinoda S, Toyoshima S, Nakazawa H, Makino T, Nakajin S . Effects of flavonoid phytochemicals on cortisol production and on activities of steroidogenic enzymes in human adrenocortical H295R cells. J Steroid Biochem Mol Biol 2002; 80 (30): 355–363.
Acknowledgements
The technical assistance provided by Dr Shiow-Chwen Tsai is appreciated. This study was supported by three grants (No. DOH92-TD-1002, No. DOH93-TD-1014 and DOH93-TD-F-113-053-(2)) from the Department of Health, Executive Yuan, Taiwan, ROC. The anti-P450scc antibody was kindly provided by Dr BC Chung, Institute of Molecular Biology, Academia Sinica, Taipei, Taiwan, ROC. The anti-StAR antibody was kindly provided by Dr DM Stocco, Department of Cell Biology and Biochemistry, Texas Tech University Health Science Center, Lubbock, Texas, USA.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hsia, SM., Chiang, W., Kuo, YH. et al. Downregulation of progesterone biosynthesis in rat granulosa cells by adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) bran extracts. Int J Impot Res 18, 264–274 (2006). https://doi.org/10.1038/sj.ijir.3901405
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.ijir.3901405
Keywords
This article is cited by
-
Endoplasmic reticulum stress induced by an ethanol extract of Coicis semen in Chang liver cells
BMC Complementary and Alternative Medicine (2018)
-
Effects of crude adlay hull acetone extract on corticosterone release from rat zona fasciculata-reticularis cells
Naunyn-Schmiedeberg's Archives of Pharmacology (2006)
