
Abstract
Here, we report the identification of the ubiquitin-like gene UBD as a downstream element of FOXP3 in human activated regulatory CD4+CD25hi T cells (Treg). Retroviral transduction of UBD in human allo-reactive effector CD4+ T helper (Th) cells upregulates CD25 and mediates downregulation of IL4 and IL5 expression similar to overexpression of FOXP3. Moreover, UBD impairs Th cell proliferation without upregulation of FOXP3 and impairs calcium mobilization. In the presence of ionomycin, overexpression of UBD in Th cells leads to the induction of IL1R2 that resemble FOXP3-transduced Th cells and naturally derived Treg cells. A comparison of the transcriptome of FOXP3- and UBD-transduced Th cells with Treg cells allowed the identification of the gene LGALS3. However, high levels of LGALS3 protein expression were observed only in human CD4+CD25hi derived Treg cells and FOXP3-transduced Th cells, whereas little was induced in UBD-transduced Th cells. Thus, UBD contributes to the anergic phenotype of human regulatory T cells and acts downstream in FOXP3 induced regulatory signaling pathways, including regulation of LGALS3 expression. High levels of LGALS3 expression represent a FOXP3-signature of human antigen-stimulated CD4+CD25hi derived regulatory T cells.
Similar content being viewed by others


Main
Regulatory CD4+CD25+ T (Treg) cells have emerged as a unique population of suppressor T cells that are important for the maintenance of peripheral tolerance.1, 2 In humans, Treg cells are characterized by high levels of CD25 expression (CD25hi) and represent 1–3% of all CD4+ T cells in peripheral blood.3 Typically, such Treg cells are anergic upon stimulation via their T-cell receptor and exhibit suppressive activity towards effector T-cell function.2 The clinical relevance of CD4+CD25hi Treg cells has recently been illustrated in different human disease settings, that is, altered numbers and impairment of suppressor activity of Treg cells has been demonstrated in patients with Hodgkin's lymphomas, autoimmune polyglandular syndrome type II, multiple sclerosis, myasthenia gravis, diabetes, psoriasis, patients with graft-versus-host-disease, and infectious diseases such as malaria and HIV.4, 5, 6, 7, 8, 9, 10, 11, 12
Development and function of Treg cells critically depend on the forkhead transcription factor FOXP3 that specifies the regulatory T cell lineage.13 As a result, humans and mice deficient in FOXP3 lack CD4+CD25+ Treg cells and suffer from multiorgan autoimmune disease.2
Treg cells normally develop in the thymus. However, they also may arise in a FOXP3-dependent manner from peripheral CD4+CD25− T cells.14, 15 The antigen-specificity of Treg cells is currently a matter of debate, although self-antigens most likely represent the dominating specificity.16, 17, 18, 19
Mechanisms of immunosuppression by Treg cells comprise cell-contact dependent suppression, mainly observed in vitro, and cytokine-mediated suppression of effector T cells, mainly observed in vivo.20 Cytokines most commonly found are IL10 and TGF-β, which also modulate Treg cell function in vivo and Treg cell development from nonregulatory CD4+CD25− T cells in vitro.14, 17, 21, 22 However, IL10 and TGF-β are not essential for the development and function of Treg cells.23, 24
CTLA4 also functions as an important mediator and modulator of Treg cell activity.20 This member of the CD28-family is constitutively expressed at high levels in CD4+CD25+ Treg cells in mouse and man. In the human system, CTLA4+ Treg cells display a stronger suppressive activity in vitro than CTLA4− Treg cells.25 However, CTLA4 is not essential for the development and function of Treg cells, since its loss could be compensated by increased secretion of IL10 and TGF-β by CTLA4−CD4+CD25+ Treg cells.26
The conversion of murine naïve nonregulatory CD4+CD25− T cells into Treg cells by retroviral overexpression of FOXP3 underlines the master control function of this transcription factor.27, 28 Consequently, the use of FOXP3 as a tool for potential clinical Treg cell therapy has been demonstrated recently in mice. Retroviral transduction of FOXP3 in polyclonal or antigen-specific naïve CD4+CD25− T cells generated Treg cells that suppressed the development of inflammatory bowl disease, autoimmune gastritis, diabetes or contact allergy, and induced transplant tolerance in vivo.27, 29, 30, 31 Despite these encouraging results in mice, debate exists about the efficacy of FOXP3 to reprogram human naïve CD4+ T cells towards a regulatory phenotype.32, 33, 34 The most likely reason for this ambiguity arises from the fact that unlike murine naïve and effector CD4+ T helper cells, human nonregulatory CD4+ T cells do express FOXP3.35
Thus, the fundamental role of FOXP3 is well established, but target genes involved in the control of the FOXP3-dependent regulatory phenotype still require defining.36 However, precise knowledge of this, including considerations of the differences between FOXP3 in mice and men,35 is essential for the effective manipulation of regulatory T cells as immunomodulatory therapy. Hence, we compared the genetic program of human CD4+CD25hi derived Treg cells, which were expanded in vitro by allogeneic stimulation and IL2, with that of human allo-reactive effector Th cells, which were reprogrammed by retroviral transduction of FOXP3. Diubiquitin (UBD) was identified as a gene that is upregulated by FOXP3 overexpression in Th cells similar to Treg cells. UBD overexpression in Th cells demonstrated that it contributes in part to the FOXP3-dependent regulatory phenotype without upregulation of FOXP3, and led to the identification of the β-galactoside binding protein LGALS3. High levels of LGALS3 represent a FOXP3-signature of human antigen-stimulated CD4+CD25hi derived Treg cells as it is not upregulated in nonregulatory CD4+CD25− derived effector Th cells despite the expression of CD25 and FOXP3.
Materials and methods
Purification and Cultivation of Human CD4+ T Cells
CD4+ T cells were prepared from peripheral blood of healthy donors by centrifugation over Ficoll-Hypaque gradients (Biochrom AG, Berlin, Germany) and MACS isolation using the CD4+ T cell isolation kit and AutoMACS technology (Miltenyi Biotech, Bergisch Gladbach, Germany). CD4+CD25− T cells and CD4+CD25hi Treg cells were then isolated by cell sorting on a MoFlo (DakoCytomation, Fort Collins, CO, USA) to a purity of >98%. CD4+CD25hi Treg cells were stimulated once with plate-bound anti-CD3 (TR66, 1 μg/ml), soluble anti-CD28 (CD28.2, 1 μg/ml, BD), and 50 U/ml recombinant human IL2 (Proleukin, provided by P Wagner, Chiron Corporation, Emeryville, CA, USA), and thereafter weekly with irradiated allogeneic EBV-transformed B cells (LG2-EBV, provided by T Boon, LICR, Brussels, Belgium). CD4+CD25− T cells were used to establish long-term, allo-reactive effector Th cell lines against LG2-EBV cells. Culture medium was IMDM, with 10% FCS, 100 U/ml penicillin/streptomycin, and nonessential amino acids (PAA Laboratories, Linz, Austria). Human peripheral blood was obtained after informed consent according to the MHH guidelines.
Antibodies
For immunostaining PE-, FITC-, APC-, and CyChrom-conjugated Abs against CD4 (RPA-T4), CD25 (M-A251), CD28 (CD28.2), TCRαβ (T10B9.1A-31), CTLA4 (BNI3), LGALS3 (B2C10; all from BD, Bioscience, San Jose, CA, USA), and FOXP3 (PCH101, eBioscine Inc., San Diego, CA, USA) and the respective isotype controls were used. For FOXP3 staining, murine hybridoma T cell transduced with human FOXP3 or GFP were included as further control. Anti-CD3ɛ (TR66, produced from hybridoma supernatants) and anti-CD28 (CD28.2, BD) were used for T-cell stimulation.
Analysing T-Cell Functions
T-cell proliferation and suppressor activity was assessed by stimulating 3 × 104 T cells in triplicate with irradiated LG2-EBV B cells or 1 μg/ml plate-bound anti-CD3 (TR66) with or without IL2, or 1 μM ionomycin (Sigma, St Louis, MO, USA), in 96-well flat-bottom microtiter plates (Nunc, Wiesbaden, Germany). For transwell experiments, Th cells were stimulated in 96 flat-bottom plates separated by 0.2 μm-pore transwell inserts (Greiner bio-one, Frickenhausen, Germany) from the T cells above the transwell. Cells were pulsed with 1 μCi/well of [3H]thymidine after 72 h for the final 16 h. Statistical analysis was performed using two-sided Student'st-test as indicated. TCR downmodulation upon anti-CD3 stimulation (plate-bound, 1 μg/ml) was assessed by FACS analysis (FACS Calibur or FACS Scan, BD) using PE-coupled anti-TCR antibodies.
Cytokine Detection
Stimulated cytokine release of IFNγ and IL10 was detected using ELISA sets of the CytoSet series (IFNγ, IL10; Biosource, Camarillo, CA, USA) and OptEIA series (IFNγ, and IL10, BD), according to the manufacturer's instructions. Supernatants of stimulated T cells were harvested at 72 h after stimulation immediately before the [3H]thymidine pulse.
Calcium Mobilization Assays
Calcium mobilization was measured by flow cytometry and confocal microscopy. In brief, T cells were loaded with 2 μM INDO-1 (Molecular Probes, Eugene, OR, USA) for 30–45 min at 37°C, extensively washed, and loaded with 1 μg/ml anti-CD3 (TR66) for 20 min at 4°C. Samples were applied to a MoFlo for 30 to 60 s at 37°C to measure the basal intracellular calcium concentration. Cells were stimulated by crosslinking cell-bound CD3 antibodies with a 1/30 dilution of goat-anti-mouse IgG antiserum or 10 μg/ml goat-anti-mouse IgGγ (Biomol International LP, Plymouth Meeting, PA, USA). The time-dependent intracellular Ca2+ was measured as the ratio of calcium-bound (405 nm) to calcium-free (515 nm) INDO-1 fluorescence, analyzed using FlowJo software (Tree Star, San Carlos, CA, USA). For single cell analysis, INDO-1 labeled cells (1 μM) were stimulated in coverslip-chambers (Nunc) precoated with 10 μg/ml anti-CD3 (TR66) using confocal microscopy (BIO-Rad, Hercules, CA, USA).
Retroviral Transduction of Human Effector CD4+ T Cells
The cDNAs encoding human FOXP3, CTLA4, and UBD were amplified from cDNA of Treg cells using high fidelity PFU polymerase (Promega) and specific primers (FOXP3: 5′-GAC AAG GAC CCG ATG CCC A-3′ and 5′-TCA GGG GCC AGG TGT AGG GT-3′; CTLA4: 5′-TAC ATA TCT GGG ATC AAA GC-3′ and 5′-ATT TGG GTT CCG CAT CCA-3′; UBD: 5′-TGC AGA GAT GGC TCC CAA TG-3′ and 5′-GAC CCC TGC CAA CAC CCC AT-3′). PCR products were cloned into pCR4.1 TOPO (Invitrogen, Carlsbad, CA, USA), sequenced, and inserted into a pMSCV-based retroviral vector encoding an enhanced GFP (GFP) under the control of an IRES sequence.37 The amphotropic PT67 packaging cell line (provided by M Wirth, GBF) was transfected as described elsewhere.37 Filtered (0.45 μm) virus-containing supernatant supplemented with 8 mg/ml sequabrene (Sigma) was applied to differentiated allo-reactive Th cells, established as described above, at day 2 after allogeneic stimulation by centrifugation at 5000 g for 60 min at room temperature. Cells were expanded thereafter with 50 U/ml IL2, and GFP expressing cells were sorted 1 to 2 weeks later using a FACS-Vantage (BD). Sorted cells were kept for up to 3 months using the restimulation procedure described above and were tested repeatedly for stable functional, phenotypic, and transcriptional changes.
Quantitative and Semiquantitative RT-PCR
Total RNA was isolated from CD4+ T cells using RNAeasy (Qiagen, Hilden, Germany) or nucleospin RNA-II (Macherey Nagel, Düren, Germany). cDNA synthesis was performed using oligo-dT primers and Superscript II Reverse Transcriptase (Invitrogen). Quantitative Real-time PCR was performed on an ABI PRISM 7000 cycler (Applied Biosystems, Foster City, CA, USA) using the SYBR Green PCR kit (Stratagene, La Jolla, CA, USA), and primers specific for FOXP3 (5′-GAA CGC CAT CCG CCA CAA CCT GA-3′ and 5′-CCC TGC CCC CAC CAC CTC TGC-3′), LGALS3 (5′-CGG AGC ACC TGC ACC TGG AGT CTA-3′ and 5′-TGA AGC GTG GGT TAA AGT GGA AGG-3′), and RPS9 (5′-CGC AGG CGC AGA CGG TGG AAG C-3′ and 5′-CGA AGG GTC TCC GCG GGG TCA CAT-3′) as described.37 For semiquantitative RT-PCR, three-fold dilutions of cDNA samples starting with the first dilution, normalized to the expression of RPS9 (5′-CGC AGG CGC AGA CGG TGG AAG C-3′ and 5′-CGT AGG AGA GCG CAG AGA GAA GTC A-3′) were amplified with primers for FOXP3 (5′-TCA ACG GTG GAT GCC CAC GC-3′ and 5′-TCA GGG GCC AGG TGT AGG GT-3′), UBD (5′-CCC AAT GCT TCC TGC CTC TGT G-3′ and 5′-GAC CCC TGC CAA CAC CCC AT-3′), IL4 (5′-AAC ACA ACT GAG AAG GAA ACC TTC TGC-3′ and 5′-CTC TCT CAT GAT CGT CTT TAG CCT TTC-3′), IL5 (5′-GAG GAT GCT TCT GCA TTT GAG TTT G-3′ and 5′-GTC AAT GTA TTT CTT TAT TAA GGA CAA G-3′), GATA3 (5′-ACC TGT CAG ACC ACC ACA ACC ACA C-3′ and 5′-GGA TGC CTT CCT TCT TCA TAG TCA GG-3′), IL1R2 (5′-AAA TGA CTC TGC TAG GAC GGT CCC-3′ and 5′-TCT TCC AGG GCC ACA TCG TG-3′) and LGALS3 (5′-CCC ATC TTC TGG ACA GCC AAG TGC-3′ and 5′-GCA TCA TTC ACT GCA ACC-3′).
GeneChip Assays and Microarray Data Analysis
For the screening of similarly regulated genes in Treg versus Th cells compared to ThFOXP3 versus ThGFP cells, we analyzed two independent experiments using Human Genome U133 A arrays (Affymetrix, Santa Clara, CA, USA) and MAS 5.0 software (www.affymetrix.com/support/technical/manuals.affx: Microarray Suite User's Guide, Version 5.0). Selection criteria for potential FOXP3-dependent genes in Treg and ThFOXP3 cells were: (1) a signal with a fold-change of more than 3 in all four comparisons, or (2) a similar increase (I, including marginal increase, MI) or decrease (D, including marginal decrease, MD) of the signal change in all four experiments. GeneChip analysis of ThUBD compared to ThGFP cells was performed using Human Genome U133 PLUS 2.0 arrays (Affymetrix). All data from ThFOXP3, ThUBD, and Treg cells were compared to each other, according to the criteria (1) a signal fold-change of more than 2, or (2) a similar signal change as either I and MD or D and MD in all six comparisons. Selected differences of one cell type were identified by (1) a signal fold-change of less than 1, and (2) an opposite signal change or no signal change (NC) in the respective T-cell population. All microarray data have been deposited at GEO (http://www.ncbi.nlm.nih.gov/geo/; GEO accession GSE4527). All T cells used for GeneChip analysis have been established for more than 2 to 3 months.
Results
Characterization of Human CD4+CD25hi Derived Treg Cells
For the generation and differentiation of human allo-reactive Th cells and Treg cells, sorted CD4+CD25− and CD4+CD25hi T cells were stimulated with allogeneic EBV-transformed B cells and IL2. CD4+CD25hi derived Treg cells expanded by this protocol could be grown for a long period of time (up to 6 months), but remained anergic to anti-CD3 stimulation when IL2 was omitted (Supplementary Figure 1a). These Treg cells consistently suppressed proliferation of established allo-reactive Th cell lines activated by anti-CD3 (Supplementary Figure 1a) or allogeneic EBV-transformed B cells (Supplementary Figure 1b). The suppressive activity of such Treg cells was overcome by the addition of more than 10 U/ml IL2 (Supplementary Figure 1b) and required cell-contact since it was blocked by a transwell membrane (Supplementary Figure 1c). Phenotypical analysis revealed stable expression of high levels of CD25, CD28, and intracellular CTLA4 (data not shown) as well as FOXP3 mRNA independent of their activation status (Supplementary Figure 1d). Thus, CD4+CD25hi derived Treg cells sorted and expanded by our protocol maintained high level of sustained FOXP3 expression associated with phenotypic and functional stability.
Ca2+ metabolism is essential for the regulation of anergy38 and has recently been shown to be impaired in murine CD4+CD25+ Treg cells analyzed ex vivo.39 In contrast to the latter report, our human Treg cells showed signs of sustained Ca2+-flux upon anti-CD3 stimulation. This was reflected in the prolonged time required to reach the lower plateau level, following the initial Ca2+-rise, and was also higher than that found for Th cells (Supplementary Figure 2a). Consistent results were obtained at the single cell level (Supplementary Figure 2b and c).
Differences between Treg and Th cells were also observed in the extent of anti-CD3 induced TCR downmodulation, which was strongly impaired in Treg cells (Supplementary Figure 2d). This finding is in line with the observation that the extent of TCR downmodulation following ligand binding correlates with the induction of effector function.40
In summary, the Treg cell lines established by our protocol showed all the phenotypic and functional characteristic of bona fide regulatory CD4+CD25hi Treg cells and thus can be used to establish the mechanisms of FOXP3-dependent gene regulation.
FOXP3 Overexpression in Human Effector Th Cells Induces a Partial Regulatory Phenotype
To identify relevant FOXP3-dependent genes associated with the regulatory phenotype, we used retroviral overexpression of FOXP3 in human allo-antigen specific effector Th cells (ThFOXP3). Successfully transduced Th cells were sorted for GFP+, reintroduced into culture, and tested repeatedly over a period of up to 3 months.
FOXP3-transduced Th cells were impaired in their proliferative response upon antigen-specific stimulation using allogeneic EBV-transformed B cells or stimulation with anti-CD3, reversed by the addition of IL2 (Figure 1a). In addition, ThFOXP3 cells acquired suppressor activity towards effector Th cells stimulated by allogeneic EBV-transformed B cells, but this was not observed in control Th cells transduced with GFP (ThGFP) or CTLA4 (ThCTLA4) (Figure 1b). Thus, FOXP3 was able to specifically reprogram differentiation of helper T cells towards a regulatory phenotype. However, compared to naturally occurring Treg cells, this regulatory phenotype was less pronounced (Figure 1a and b).

FOXP3 is able to reprogram human allo-reactive effector Th cells towards a regulatory phenotype. (a) Treg cells and Th cells, retrovirally transduced with FOXP3 plus GFP (ThFOXP3), CTLA4 and GFP (ThCTLA4), and GFP alone (ThGFP), were stimulated with allogeneic LG2-EBV B cells or anti-CD3 without or with 100 U/ml IL2. Proliferation was assessed at day 3. (b) ThGFP cells as in (a) were stimulated by allogeneic LG2-EBV B cells in the presence of equal numbers of indicated T cells. Only Treg and ThFOXP3 cells significantly suppressed ThGFP cell proliferation; NS=not significant. Similar results were obtained with normal effector Th cells instead of ThGFP cells. (c) Th cells as in (a) were tested for intracellular CTLA4 expression in flow cytometry. Shaded area: isotype control, bold line: intracellular anti-CTLA4 staining. (d) Transduced Th cells as in (a) were tested for downmodulation of TCR expression following anti-CD3 stimulation at the times indicated. Remaining surface TCR expression was measured with anti-TCRαβ antibodies by flow cytometry (MFI: mean fluorescence intensity). (e) Sustained overexpression of FOXP3 in ThFOXP3 compared to Th GFP cells. Real-time RT-PCR was carried out at the times indicated after stimulation with plate-bound anti-CD3 plus IL2. (f) Intranuclear FOXP3 protein expression of ThFOXP3 cells compared to Treg and ThGFP cells was measured by flow cytometry. Gates were set according to the control staining using murine hybridoma T cells transduced with GFP alone (negative control; gray shaded) or with human FOXP3 plus GFP (thin line). Percent of positive cells is indicated.
In contrast, overexpression of CTLA4 did not induce a regulatory phenotype in Th cells, although the levels of intracellular CTLA4 expression were similar to Treg cells (Figure 1c). A possible contribution of CTLA4 to the FOXP3-dependent transcriptional control is therefore unresolved at this point.
We then analyzed the anti-CD3 induced TCR downmodulation. Similar to Treg cells, ThFOXP3 cells showed an impaired downmodulation of TCR expression that was not observed in ThGFP or ThCTLA4 cells (Figure 1d). Interestingly, this impairment of TCR downmodulation appears to be a general effect of FOXP3 as it was even observed in murine hybridoma T cells transduced with FOXP3. However, such hybridoma T cells did not acquire an anergic phenotype, although they showed upregulation of CD25 and impaired NFAT function (data not shown). Therefore, a high level of sustained FOXP3 mRNA (Figure 1e) and protein expression in ThFOXP3 cells (Figure 1f) induced most of the characteristics of the FOXP3-dependent regulatory phenotype.
It should be noted that in long-term cultures, ThFOXP3 cells were able to escape the FOXP3-dependent control. Most likely, the appearance of T cells with lower expression levels of FOXP3 was responsible for this phenomenon, since such T cells also showed reduced expression of the co-transduced IRES-driven GFP. As a result, re-sorting of long-term cultured ThFOXP3 cells for high expression of GFP re-established their regulatory phenotype (data not shown).
In addition, ThFOXP3 cells did not reach the same high levels of CD25 expression (Figure 2a) while CD28 was present at similar levels in both types of cells (data not shown). We, therefore, analyzed the functional properties of ThFOXP3 cells expressing different levels of CD25 (Figure 2b). CD25hi ThFOXP3 cells showed a more pronounced regulatory phenotype than CD25lo ThFOXP3 cells in terms of their proliferative impairment and suppressor function (Figure 2c and d). Again, CD25 expression decreased in long-term culture despite the presence of FOXP3 (data not shown). This suggests that the Th cell specific regulation of CD25 expression is only transiently overcome by FOXP3 overexpression in differentiated effector Th cells.

Differences in the regulatory phenotype between CD25hi and CD25lo Th cells transduced with FOXP3. (a) Th cells transduced with FOXP3 plus GFP (ThFOXP3) and GFP alone (ThGFP) were tested for CD25 expression and compared with Treg and Th cells. FOXP3 transduction never resulted in CD25 expression comparable to bona fide Treg cells. (b) Reanalysis of ThFOXP3 cells sorted for CD25hi and CD25lo expression is shown. (c) ThFOXP3 cells sorted for CD25hi and CD25lo expression, Th and ThGFP cells were stimulated with allogeneic LG2-EBV B cells with or without 100 U/ml IL2. CD25hi ThFOXP3 cells are more impaired in their proliferative capacity than CD25lo ThFOXP3 cells. (d) ThFOXP3 cells of (c) were added to Th cells in equal numbers and stimulated with allogeneic LG2-EBV B cells. Suppressive activity was significantly more pronounced (*P<0.02) in CD25hi ThFOXP3 than CD25lo ThFOXP3 cells.
Gene Expression Profiles of Treg Cells and ThFOXP3 Cells
The data presented above provide a biological framework for the identification of genes associated with the FOXP3-dependent regulatory phenotype. Using GeneChip analysis, we screened for genes similarly regulated in both Treg and ThFOXP3 cells compared to Th and ThGFP cells. According to our stringent selection criteria, 39 genes were defined that were similarly regulated in both regulatory T-cell populations. As expected, known markers of Treg cells were identified, that is CD25, CTLA4, TNFRSF1B, and CCR7. In addition, we found genes that were not previously associated with FOXP3-dependent transcriptional control, for instance the ubiquitin-like gene diubiquitin (UBD) and the β-galactoside binding lectin LGALS3 (Table 1). Proteins involved in ubiquitinylation play an important role in the regulation of T-cell anergy.41 Hence, we analyzed the potential contribution of UBD to the FOXP3-dependent transcriptional control in more detail.
UBD is Involved in the Regulation of CD4+ T-Cell Anergy
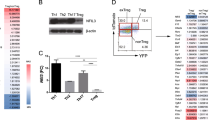
UBD is expressed at high levels in CD4+CD25hi derived Treg cells and consistently upregulated in FOXP3-overexpressing Th cells compared to ThGFP and ThCTLA4 cells (Figure 3a). Unlike FOXP3, which is expressed at sustained high levels, UBD expression is transiently downregulated following T-cell activation using anti-CD3/-CD28 antibodies and IL2 (Figure 3a).

UBD contributes to the transcriptional and functional control of FOXP3. (a) Th cells transduced as in Figure 1a were tested for UBD mRNA expression by semiquantitative RT-PCR after stimulation with plate-bound anti-CD3, soluble anti-CD28, and 100 U/ml IL2. cDNA was tested in three-fold dilutions starting with the first dilution; RPS9 was used as house-keeping control. (b) Th cells transduced with UBD plus GFP (ThUBD), FOXP3 plus GFP (ThFOXP3), and GFP alone (ThGFP) were tested for surface CD25 and CD28 expression in flow cytometry. (c) Treg cells and Th cells as in b were stimulated with LG2-EBV B cells alone or in the presence of 100 U/ml IL2. Proliferation was assessed at day 3. (d) The same T cells as in c were analyzed for IFNγ and IL10 cytokine secretion following stimulation with LG2-EBV B cells.
Overexpression of UBD in Th cells (ThUBD) via retroviral transduction induced upregulation of CD25 without affecting CD28 expression (Figure 3b). In long-term ThUBD cell lines, significant impairment of proliferation upon TCR stimulation was observed, which was reversed by the addition of IL2 (Figure 3c). Similar to FOXP3, overexpression of UBD in effector Th cells did not suppress IL10 and IFNγ secretion (Figure 3d). Importantly, FOXP3 expression was not upregulated in ThUBD cells (Figure 4a and b) suggesting that UBD is a key player in the regulation of anergy in Treg cells downstream of FOXP3.

UBD expression impairs T-cell proliferation independent of FOXP3. (a) Effector Th cells transduced as described in Figure 3b were tested by semiquantitative RT-PCR as described in Figure 3a for mRNA of genes known to be regulated in FOXP3 transduced Th cells. Note that FOXP3 is not upregulated in ThUBD cells. (b) Expression of high levels of FOXP3 protein in ThFOXP3 cells, but not in ThGFP and ThUBD cells, determined by flow cytometry as described in Figure 1f. Percent of positive cells is indicated. (c) Quantification of LGALS3 mRNA by Real-time RT-PCR in Treg and ThFOXP3 cells compared to ThGFP cells. (d) Anti-CD3 induced Ca2+-mobilization in ThFOXP3, ThUBD, and ThGFP cells was determined on a MoFlo as described in Supplementary Figure 2a. (e) ThGFP and ThUBD cells were tested for upregulation of IL1R2, FOXP3, and UBD mRNA. Ionomycin (Io) or vehicle (DMSO) was added. Determination was carried out after 24 h by semiquantitative RT-PCR as described in Figure 3a. (f) IL1R2 specific mRNA was determined in ThFOXP3, ThUBD, and ThGFP cells of Figure 3b by semiquantitative RT-PCR. Note that no signal is obtained in ThUBD as in ThGFP cells. The slight discrepancy between e and f in this respect is most likely due to differences between the individual donors of Th cells.
Like ThFOXP3 cells, mRNA of IL5 and IL4 was significantly downregulated in ThUBD cells (Figure 4a). Whereas GATA3, regulating Th2 specific IL5 and IL4 cytokine expression, was unaltered (Figure 4a),42 LGALS3, which has been described to downregulate expression of IL4 and IL5 mRNA,43 was only slightly induced (Figure 4a and c). Thus, UBD appears to be responsible for the IL4/IL5 downregulation as a downstream effector of FOXP3.
We then analyzed the influence of UBD overexpression on TCR downmodulation and Ca2+-mobilization. Interestingly, UBD did not impair the anti-CD3 induced TCR downmodulation observed with Treg cells and ThFOXP3 cells (data not shown). However, Ca2+-mobilization was influenced (Figure 4d). Whereas FOXP3 overexpression induced a slightly elevated steady-state phase of Ca2+-flux compared to ThGFP cells, this state was lower in Th cells overexpressing UBD (Figure 4d).
Since ionomycin is known to induce ubiuqitin-ligases associated with anergy,38 we investigated the effect of ionomycin treatment on UBD expression in Th cells. As predicted, ionomycin induced upregulation of UBD mRNA (Figure 4e), but at the same time, FOXP3 mRNA expression was downregulated despite overexpression of UBD (Figure 4e). This suggests that the ionomycin-induced anergic state is independent of FOXP3 and confirms that UBD does not induce the expression of FOXP3. Interestingly, similar to overexpression of FOXP3 in Th cells (Figure 4f), overexpression of UBD together with ionomycin-induced sustained Ca2+-flux upregulated IL1R2 mRNA expression (Figure 4e). As FOXP3 overexpression leads to the upregulation of UBD and in parallel increases the levels of TCR-stimulated Ca2+-flux, both UBD together with an elevated Ca2+-flux seem to cooperatively regulate the expression of IL1R2. On the other hand, ionomycin did not improve the suppressive activity of Th cells overexpressing UBD (data not shown), indicating that high levels of FOXP3 are required to obtain a suppressive phenotype.
High LGALS3 Protein Expression Specifies a FOXP3-Dependent Signature of Human Treg Cells
We extended our GeneChip expression analysis of ThFOXP3 and Treg cells to ThUBD cells. Regulated genes could be classified into four groups (Figure 5a). (i) Genes regulated in all three T-cell populations. These included UBD and LGALS3. (ii) Genes expressed similarly in ThFOXP3 and ThUBD cells only, such as CD9. (iii) Genes expressed similarly in ThUBD and Treg cells only, such as CBL-B. (iv) Genes expressed similarly in ThFOXP3 and Treg cells only, such as CCR7, FYB, and CCNG2. These data reflect the congruencies and differences between naturally derived Treg cells, and our FOXP3 and UBD engineered effector Th cells, respectively.

LGALS3 protein expression represents a FOXP3-signature of CD4+CD25hi derived Treg cells. (a) Transcriptome analysis of ThUBD, ThFOXP3, and Treg cells. Four groups were defined when analyzing significantly regulated genes. ThUBD=ThFOXP3=Treg: genes regulated similarly in all three regulatory types of T cells including UBD and LGALS3. ThUBD=Treg: genes regulated similarly in regulatory T cells and UBD transduced Th cells. ThUBD=ThFOXP3: genes regulated similarly in UBD and FOXP3 transduced Th cells. ThFOXP3=Treg: genes regulated similarly in regulatory T cells and FOXP3 transduced Th cells. The signal fold change is represented as red (upregulated) or green color (downregulated) indicated below the panels. (b) Treg and Th cells were tested for intracellular LGALS3 expression in flow cytometry. Thin line: isotype control; bold line: anti-LGALS3 staining. (c) LGALS3 expression is not induced following T-cell stimulation. Th and Treg cells were tested for the expression of FOXP3, CD25, LGALS3, and CTLA4 at day 3 following allogeneic stimulation using LG2-EBV B cells and IL2. (d) Specificity of FOXP3 staining was confirmed using murine hybridoma T cells transduced with GFP alone (GFP) and human FOXP3 and GFP (FOXP3) as described in Figure 1f. Shaded area: isotype control, bold line: antibody staining. Percent of positive cells is indicated. (e) Surface staining of CD4 and CD25 on human peripheral blood lymphocytes of a healthy donor (left panel). Regions and percentage of CD4+ T cells expressing high, intermediary, and no CD25 expression, respectively, correspond to the colors in the right panel (CD25hi=red, CD25int=green, CD25neg=blue curve), which depicts the corresponding intracellular LGALS3 staining. (f) Th cells were stimulated by allogeneic LG2-EBV B cells in the presence of Treg cells at the indicated ratios with or without 50 mM lactose. Proliferation was assessed at day 3.
Since upregulation of LGALS3 mRNA was common to Treg cells and Th cells overexpressing FOXP3 and UBD, respectively, we analyzed LGALS3 protein expression in these cells. Interestingly, high sustained levels of LGALS3 protein expression were detected only in Treg cells (Figure 5b). Whereas significant induction of LGALS3 protein was also observed in ThFOXP3 cells, UBD overexpression in Th cells only minimally affected LGALS3 protein expression (Figure 5b). Contrary to mice,44 LGALS3 protein was not significantly induced in human nonregulatory effector T cells following stimulation with allogeneic EBV-transformed B cells and IL2, although upregulation of intranuclear FOXP3, cell surface CD25, and intracellular CTLA4 protein expression was observed in such cells (Figure 5c). As LGALS3 protein expression is restricted at low levels to nonactivated human peripheral blood CD4+CD25+ T cells (Figure 5e), high level of LGALS3 protein expression represents a Treg-specific FOXP3 signature of antigen-stimulated human CD4+CD25hi-derived Treg cells.
LGALS3 has been described to induce T-cell apoptosis in human T cells via a lactose-dependent binding to CD7 and CD29.45 Therefore, we analyzed the suppressor function of Treg cells in the presence or absence of 50 mM lactose, which potently impairs the apoptotic function of LGALS3.45 But no impairment of Treg cell suppressor function was observed suggesting that LGALS3 is not directly involved in Treg cell suppression (Figure 5f).
Discussion
In the present study we compared the genetic program of human CD4+CD25hi derived Treg cells with that of allo-reactive effector Th cells reprogrammed towards a partial regulatory phenotype by retroviral transduction of FOXP3. The reason for such an approach was to unmask FOXP3-dependent genes by mimicking sustained high levels of FOXP3 expression in Th cells and to reduce the size of differentially expressed genes to the most significant set. Critical FOXP3-dependent target genes were identified.
One of the genes consistently upregulated was the ubiquitin-like gene diubiquitin (UBD, also known as FAT10). UBD has been initially cloned as a gene exclusively expressed in dendritic cells and mature B cells.46 It was reported to be involved in protein degradation, apoptosis, and control of spindle assembly.47, 48 Moreover, UBD expression can be induced by IFNγ, whereas another IFNγ inducible gene, NEDD8 ultimate buster-1L (NUB1L), has been reported to accelerate the degradation of UBD.49 A potential function of UBD in the physiology of human Treg cells has not been addressed so far. This is most likely due to the fact that UBD expression is found at significant levels only after in vitro activation as demonstrated in this work.
Since FOXP3 is a transcriptional repressor,36, 50 upregulation of UBD expression should not be directly induced by FOXP3. Rather, UBD transcription should be indirectly regulated, for example, by downregulating transcriptional repressors of UBD. This is supported by preliminary experiments in which a 1.5 kb genomic DNA fragment preceding the transcriptional start of UBD was tested in a reporter system. No reporter activity was induced by FOXP3 (unpublished data).
The contribution of high levels of UBD expression to the FOXP3-dependent regulatory phenotype became apparent, as we could show that UBD overexpression in Th cells interfered with proliferation and upregulated cell surface expression of CD25. In addition, expression of IL4 and IL5 mRNA was downregulated in ThUBD cells comparable to Th cells overexpressing FOXP3, although FOXP3 was not induced by UBD. In contrast, genes normally induced upon FOXP3-overexpression, for example CD28 and CTLA4, were not affected by UBD transduction. This indicates that UBD acts downstream in some of the FOXP3-induced regulatory signaling branches, and, therefore, only a partial regulatory phenotype is induced by UBD alone as it does not upregulate FOXP3. A clear confirmation of this phenomenon was obtained, when ionomycin was added to Th cells overexpressing UBD. Only the ionomycin-mediated higher Ca2+-levels together with overexpression of UBD mimicked the FOXP3-induced transcriptional control of IL1R2. In line with this was the observation that UBD overexpression only minimally induced LGALS3 protein expression in Th cells, although LGALS3 mRNA was upregulated similar to Th cells overexpressing FOXP3.
How UBD mediates transcriptional control is questionable. It is known that E3-ubiquitin ligases such as Itch, Cbl-b, and Grail play an essential role in the induction of T-cell anergy. Their mode of action is to target key TCR signaling molecules for degradation that are localized in the cytoplasm.38, 51 UBD has similarly been reported to target proteins for degradation but not in the context of TCR signal transduction. However, UBD has also been found to be localized in the nucleus.52 Therefore, its main effect might be a transcriptional modulation, as the activity of many signal transduction pathways and transcription factors are regulated by ubiquitin-mediated proteolytic and nonproteolytic mechanisms.53, 54
Whatever the mechanisms, our extended transcriptome analysis of Treg cells compared to ThFOXP3 and ThUBD cells suggested that LGALS3 might play a central role for some of the overlapping functional and phenotypic changes observed in these cells.
LGALS3 represents a unique member of the family of β-galactoside-binding proteins. It is composed of a small N-terminal domain, which controls its subcellular targeting, a repetitive collagen-like region that serves as substrate for matrix metalloproteinases, and a C-terminal carbohydrate-recognition domain.55 Expression of LGALS3 has been observed in different tissues and cells types. It displays pleiotropic biological functions depending on subcellular localization. Extracellular LGALS3 binds to cells through glycosylated receptors, thereby triggering or modulating cellular responses such as mediator release, apoptosis, cell adhesion, and migration.45, 56 Intracellular LGALS3 has been reported to inhibit apoptosis57 and the cell cycle.58 Interestingly, upregulation of LGALS3 has been observed during neoplastic transformation in several human malignancies.59 Moreover, LGALS3 has been found in the nucleus, where it is reported to be involved in pre-mRNA splicing,60 and the regulation of gene expression (eg cyclinD and Muc2) through the activation of specific transcription factors, including AP1, NFAT, SP1, and CREB.61, 62 Serine phosphorylation and dephosporylation at position 6 of LGALS3 by casein kinase 1 and protein phosphatase 1, respectively, controls the carbohydrate binding capability, nuclear localization, transcriptional modulation, and antiapoptotic function of LGALS3.58, 63, 64 Diversified functions of LGALS3 have also been reported for cells of the immune system. LGALS3 has been detected in monocytes and macrophages, dendritic cells, mast cells, murine T cells, and granulocytes.65 Several important processes and functions are modulated by LGALS3 in these cells including thymocyte migration,66 innate immunity against infections,67 inflammation,68 and T-cell apoptosis.45, 69 Owing to the non-differential expression of LGALS3 in activated murine CD8+ and CD4+ T cells,44 it was unexpected that LGALS3 might act as a Treg-specific marker. However, only low levels of LGALS3 protein were detectable in human peripheral blood CD4+CD25+ T cells, but sustained high levels were induced in naturally derived human CD4+CD25hi Treg cells following antigenic stimulation in vitro. Similarly, upregulation of LGALS3 protein expression was observed in FOXP3-transduced effector Th cells reprogrammed towards a partial regulatory phenotype. This indicates that LGALS3 protein expression in fact represents a FOXP3-signature of the regulatory program.
The potential contribution of LGALS3 to the function of activated Treg cells is speculative. The anti-apoptotic activity of LGALS3 might protect Treg cells from apoptotic signals through CD95,70 which is expressed at high levels on Treg cells. Alternatively, the activation and modulation of specific transcription factors, like AP1, NFAT, SP1, and CREB,61, 62 might contribute to the FOXP3-dependent transcriptional control. However, more direct effects of LGALS3 on Treg cell functions have to be considered. In murine T cells, a direct association of LGALS3 with the TCR complex, restricting TCR signaling, has been reported. This interaction dependents on N-glycosylation mediated by the enzyme beta-1,6 N-acetylglucosaminyltransferase V (Mgat5).71 Therefore, mice deficient in Mgat5 show a lowered T-cell activation threshold with enhanced recruitment of TCRs to agonist-coated beads, and actin microfilament re-organization.71 In contrast to the latter, in human CD4+CD25hi derived Treg cells, that express LGALS3, we have observed a dynamic impairment of actin remodeling compared to CD4+CD25− T cells.72
The addition of soluble LGALS3 to Jurkat T cells has been reported to trigger a sustained influx of extracellular Ca2+ that can be blocked by lactose.73 As a sustained Ca2+ flux in Treg cells following TCR stimulation was observed in our case, LGALS3 released from the cells might act in an autocrine manner and modulate the Ca2+ homeostasis of Treg cells. However, similar to T cells of mice,44 we did not detect significant cell surface bound LGALS3 in resting or activated Treg cells. We, therefore, excluded a paracrine effect of LGALS3 in the process of suppression of antigen-induced Th cell proliferation by Treg cells, suggesting that LGALS3 released by Treg cells might not reach amounts needed to induce either apoptosis or sustained Ca2+ flux.69, 73
In general, the approach to reprogram antigen-specific effector Th cells into therapeutic regulatory T cells by the transduction of FOXP3 is very encouraging. Clearly, however, only a partial regulatory phenotype can be induced by FOXP3 overexpression in effector Th cells compared to CD4+CD25hi derived Treg cells. FOXP3-transduced Th cells show less proliferative impairment and suppressor function, and less induction of LGALS3 expression. They further downregulate CD25 expression, and still secrete effector cytokines like IFNγ. Based on the observation that FOXP3 protein is expressed in human effector Th cells, inevitable mechanisms must exist that control the overwhelming negative effects of FOXP3 in human effector CD4+ T cells.
Our analysis identified LGALS3 as a FOXP3-dependent signature differentiating between a regulatory and an effector program. LGALS3 is expressed at high constitutive levels only in antigen-stimulated CD4+CD25hi derived Treg cells. Moreover, we identified UBD as a downstream effector of FOXP3 involved mainly in the regulation of anergy. Thus, our analysis provides a focused starting point for further elucidation of genetic and mechanistic aspects of active immunosuppression and of differences between human and mouse regulatory T cells. This will ultimately lead to a complete understanding of human regulatory T cells.
Accession codes
References
Schwartz RH . Natural regulatory T cells and self-tolerance. Nat Immunol 2005;6:327–330.
Sakaguchi S . Naturally arising FOXP3-expressing CD25(+)CD4(+) regulatory T cells in immunological tolerance to self and non-self. Nat Immunol 2005;6:345–352.
Baecher-Allan C, Viglietta V, Hafler DA . Human CD4+CD25+ regulatory T cells. Semin Immunol 2004;16:89–98.
Marshall NA, Christie LE, Munro LR, et al. Immunosuppressive regulatory T cells are abundant in the reactive lymphocytes of Hodgkin lymphoma. Blood 2004;103:1755–1762.
Balandina A, Lecart S, Dartevelle P, et al. Functional defect of regulatory CD4+CD25+ T cells in the thymus of patients with autoimmune Myasthenia Gravis. Blood 2004;105:735–741.
Kriegel MA, Lohmann T, Gabler C, et al. Defective suppressor function of human CD4+ CD25+ regulatory T cells in autoimmune polyglandular syndrome type II. J Exp Med 2004;199:1285–1291.
Viglietta V, Baecher-Allan C, Weiner HL, et al. Loss of functional suppression by CD4+CD25+ regulatory T cells in patients with multiple sclerosis. J Exp Med 2004;199:971–979.
Lindley S, Dayan CM, Bishop A, et al. Defective suppressor function in CD4+CD25+ T-cells from patients with type 1 diabetes. Diabetes 2005;54:92–99.
Miura Y, Thoburn CJ, Bright EC, et al. Association of FOXP3 regulatory gene expression with graft-versus-host disease. Blood 2004;104:2187–2193.
Sugiyama H, Gyulai R, Toichi E, et al. Dysfunctional blood and target tissue CD4+CD25 high regulatory T cells in psoriasis: mechanism underlying unrestrained pathogenic effector T cell proliferation. J Immunol 2005;174:164–173.
Walther M, Tongren JE, Andrews L, et al. Upregulation of TGF-beta, FOXP3, and CD4(+)CD25(+) regulatory T cells correlates with more rapid parasite growth in human malaria infection. Immunity 2005;23:287–296.
Andersson J, Boasso A, Nilsson J, et al. Cutting edge: the prevalence of regulatory T cells in lymphoid tissue is correlated with viral load in HIV-infected patients. J Immunol 2005;174:3143–3147.
Fontenot JD, Rasmussen JP, Williams LM, et al. Regulatory T cell lineage specification by the forkhead transcription factor foxp3. Immunity 2005;22:329–341.
Fantini MC, Becker C, Monteleone G, et al. Cutting edge: TGF-{beta} induces a regulatory phenotype in CD4+CD25− T cells through FOXP3 induction and down-regulation of Smad7. J Immunol 2004;172:5149–5153.
Walker MR, Carson BD, Nepom GT, et al. De novo generation of antigen-specific CD4+CD25+ regulatory T cells from human CD4+CD25− cells. Proc Natl Acad Sci 2005;102:4103–4108.
Wang HY, Lee DA, Peng G, et al. Tumor-specific human CD4+ regulatory T cells and their ligands: implications for immunotherapy. Immunity 2004;20:107–118.
Nishikawa H, Kato T, Tawara I, et al. Definition of target antigens for naturally occurring CD4+ CD25+ regulatory T cells. J Exp Med 2005;201:681–686.
Hsieh CS, Liang Y, Tyznik AJ, et al. Recognition of the peripheral self by naturally arising CD25+ CD4+ T cell receptors. Immunity 2004;21:267–277.
Wang HY, Peng G, Guo Z, et al. Recognition of a new ARTC1 peptide ligand uniquely expressed in tumor cells by antigen-specific CD4+ regulatory T cells. J Immunol 2005;174:2661–2670.
von Boehmer H . Mechanisms of suppression by suppressor T cells. Nat Immunol 2005;6:338–344.
Zheng SG, Gray JD, Ohtsuka K, et al. Generation ex vivo of TGF-beta-producing regulatory T cells from CD4+CD25− precursors. J Immunol 2002;169:4183–4189.
Marie JC, Letterio JJ, Gavin M, et al. TGF-{beta}1 maintains suppressor function and FOXP3 expression in CD4+CD25+ regulatory T cells. J Exp Med 2005;201:1061–1067.
Piccirillo CA, Letterio JJ, Thornton AM, et al. CD4+CD25+ regulatory T cells can mediate suppressor function in the absence of transforming growth factor {beta}1 production and responsiveness. J Exp Med 2002;196:237–246.
Suri-Payer E, Cantor H . Differential cytokine requirements for regulation of autoimmune gastritis and colitis by CD4(+)CD25(+) T cells. J Autoimmun 2001;16:115–123.
Birebent B, Lorho R, Lechartier H, et al. Suppressive properties of human CD4+CD25+ regulatory T cells are dependent on CTLA4 expression. Eur J Immunol 2004;34:3485–3496.
Tang Q, Boden EK, Henriksen KJ, et al. Distinct roles of CTLA4 and TGF-beta in CD4+CD25+ regulatory T cell function. Eur J Immunol 2004;34:2996–3005.
Hori S, Nomura T, Sakaguchi S . Control of regulatory T cell development by the transcription factor FOXP3. Science 2003;299:1057–1061.
Fontenot JD, Gavin MA, Rudensky AY . FOXP3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol 2003;4:330–336.
Jaeckel E, von Boehmer H, Manns MP . Antigen-specific FoxP3-transduced T-cells can control established type 1 diabetes. Diabetes 2005;54:306–310.
Chai JG, Xue SA, Coe D, et al. Regulatory T cells, derived from naive CD4+. Transplantation 2005;79:1310–1316.
Loser K, Hansen W, Apelt J, et al. In vitro generated regulatory T cells induced by FOXP3-retrovirus infection control murine contact allergy and systemic autoimmunity. Gene Therapy 2005;12:1294–1304.
Yagi H, Nomura T, Nakamura K, et al. Crucial role of FOXP3 in the development and function of human CD25+CD4+ regulatory T cells. Int Immunol 2004;16:1643–1656.
Oswald-Richter K, Grill SM, Shariat N, et al. HIV infection of naturally occurring and genetically reprogrammed human regulatory T-cells. PLoS Biol 2004;2:955–966.
Allan SE, Passerini L, Bacchetta R, et al. The role of 2 FOXP3 isoforms in the generation of human CD4 Tregs. J Clin Invest 2005;115:3276–3284.
Ziegler SF . FOXP3: of mice and men. Ann Rev Immunol 2006;24:209–229.
Carson BD, Lopes JE, Soper DM, et al. Insights into transcriptional regulation by FOXP3. Front Biosci 2006;11:1607–1619.
Bruder D, Probst-Kepper M, Westendorf AM, et al. Frontline: Neuropilin-1: a surface marker of regulatory T cells. Eur J Immunol 2004;34:623–630.
Heissmeyer V, Macian F, Im SH, et al. Calcineurin imposes T cell unresponsiveness through targeted proteolysis of signaling proteins. Nat Immunol 2004;5:255–265.
Gavin MA, Clarke SR, Negrou E, et al. Homeostasis and anergy of CD4(+)CD25(+) suppressor T cells in vivo. Nat Immunol 2002;3:33–41.
Bachmann MF, Oxenius A, Speiser DE, et al. Peptide-induced T cell receptor downregulation on naive T cells predicts agonist/partial agonist properties and strictly correlates with T cell activation. Eur J Immunol 1997;27:2195–2203.
Mueller DL . E3 ubiquitin ligases as T cell anergy factors. Nat Immunol 2004;5:883–890.
Skapenko A, Leipe J, Niesner U, et al. GATA-3 in human T cell helper type 2 development. J Exp Med 2004;199:423–428.
Cortegano I, del Pozo V, Cardaba B, et al. Galectin-3 down-regulates IL5 gene expression on different cell types. J Immunol 1998;161:385–389.
Joo HG, Goedegebuure PS, Sadanaga N, et al. Expression and function of galectin-3, a {beta}-galactoside-binding protein in activated T lymphocytes. J Leukoc Biol 2001;69:555–564.
Fukumori T, Takenaka Y, Yoshii T, et al. CD29 and CD7 mediate galectin-3-induced type II T-cell apoptosis. Cancer Res 2003;63:8302–8311.
Bates EE, Ravel O, Dieu MC, et al. Identification and analysis of a novel member of the ubiquitin family expressed in dendritic cells and mature B cells. Eur J Immunol 1997;27:2471–2477.
Liu YC, Pan J, Zhang C, et al. A MHC-encoded ubiquitin-like protein (FAT10) binds noncovalently to the spindle assembly checkpoint protein MAD2. Proc Natl Acad Sci 1999;96:4313–4318.
Raasi S, Schmidtke G, Groettrup M . The ubiquitin-like protein FAT10 forms covalent conjugates and induces apoptosis. J Biol Chem 2001;276:35334–35343.
Hipp MS, Raasi S, Groettrup M, et al. NEDD8 ultimate buster-1L interacts with the ubiquitin-like protein FAT10 and accelerates its degradation. J Biol Chem 2004;279:16503–16510.
Schubert LA, Jeffery E, Zhang Y, et al. Scurfin (FOXP3) acts as a repressor of transcription and regulates T cell activation. J Biol Chem 2001;276:37672–37679.
Soares L, Seroogy C, Skrenta H, et al. Two isoforms of otubain 1 regulate T cell anergy via GRAIL. Nat Immunol 2004;5:45–54.
Lee CG, Ren J, Cheong IS, et al. Expression of the FAT10 gene is highly upregulated in hepatocellular carcinoma and other gastrointestinal and gynecological cancers. Oncogene 2003;22:2592–2603.
Conaway RC, Brower CS, Conaway JW . Emerging roles of ubiquitin in transcription regulation. Science 2002;296:1254–1258.
Gao M, Karin M . Regulating the regulators: control of protein ubiquitination and ubiquitin-like modifications by extracellular stimuli. Mol Cell 2005;19:581–593.
Barondes SH, Cooper DN, Gitt MA, et al. Galectins. Structure and function of a large family of animal lectins. J Biol Chem 1994;269:20807–20810.
Ochieng J, Furtak V, Lukyanov P . Extracellular functions of galectin-3. Glycoconj J 2004;19:527–535.
Matarrese P, Tinari N, Semeraro ML, et al. Galectin-3 overexpression protects from cell damage and death by influencing mitochondrial homeostasis. FEBS Lett 2000;473:311–315.
Yoshii T, Fukumori T, Honjo Y, et al. Galectin-3 phosphorylation is required for its anti-apoptotic function and cell cycle arrest. J Biol Chem 2002;277:6852–6857.
Takenaka Y, Fukumori T, Raz A . Galectin-3 and metastasis. Glycoconj J 2004;19:543–549.
Park JW, Voss PG, Grabski S, et al. Association of galectin-1 and galectin-3 with Gemin4 in complexes containing the SMN protein. Nucl Acids Res 2001;29:3595–3602.
Lin HM, Pestell RG, Raz A, et al. Galectin-3 enhances cyclin D(1) promoter activity through SP1 and a cAMP-responsive element in human breast epithelial cells. Oncogene 2002;21:8001–8010.
Walzel H, Blach M, Hirabayashi J, et al. Galectin-induced activation of the transcription factors NFAT and AP-1 in human Jurkat T-lymphocytes. Cell Signal 2002;14:861–868.
Mazurek N, Conklin J, Byrd JC, et al. Phosphorylation of the beta-galactoside-binding protein galectin-3 modulates binding to its ligands. J Biol Chem 2000;275:36311–36315.
Takenaka Y, Fukumori T, Yoshii T, et al. Nuclear export of phosphorylated Galectin-3 regulates its antiapoptotic activity in response to chemotherapeutic drugs. Mol Cell Biol 2004;24:4395–4406.
Chen H, Liu F, Yang R . Roles of galectin-3 in immune responses. Arch Immunol Ther Exp 2005;53:497–504.
Savino W, Mendes-da-Cruz DA, Smaniotto S, et al. Molecular mechanisms governing thymocyte migration: combined role of chemokines and extracellular matrix. J Leukoc Biol 2004;75:951–961.
Sato S, Nieminen J . Seeing strangers or announcing the the ‘danger’: Galectin-3 in two models of innate immunity. Glycoconj J 2002;19:583–591.
Rubinstein N, Ilarregui JM, Toscano MA, et al. The role of galectins in the initiation, amplification and resolution of the inflammatory response. Tissue Antigens 2004;64:1–12.
Stillman BN, Hsu DK, Pang M, et al. Galectin-3 and Galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J Immunol 2006;176:778–789.
Fukumori T, Takenaka Y, Oka N, et al. Endogenous Galectin-3 determines the routing of CD95 apoptotic signaling pathways. Cancer Res 2004;64:3376–3379.
Demetriou M, Granovsky M, Quaggin S, et al. Negative regulation of T-cell activation and autoimmunity by Mgat5N-glycosylation. Nature 2001;409:733–739.
Sechi AS, Buer J, Wehland J, et al. Changes in actin dynamics at the T-cell/APC interface: implications for T-cell anergy? Immunol Rev 2002;189:98–110.
Dong S, Hughes RC . Galectin-3 stimulates uptake of extracellular Ca2+ in human Jurkat T-cells. FEBS Lett 1996;395:165–169.
Acknowledgements
We thank Maria Höxter, Tanja Töpfer, Karen Wagner, Hanne Herrmann, and Patricia Gatzlaff for excellent technical support. We gratefully acknowledge Dr H Hauser for discussion and support, and Dr V Wray for critical reading. This work was supported by grants from the VolkswagenStiftung (I/73 234) and the Deutsche Forschungsgemeinschaft (PR 554/2, SFB621).
Author information
Authors and Affiliations
Corresponding author
Additional information
Duality of interest
None declared.
Supplementary Information accompanies the paper on the Laboratory Investigation website (http://www.nature.com/labinvest).
Rights and permissions
About this article
Cite this article
Ocklenburg, F., Moharregh-Khiabani, D., Geffers, R. et al. UBD, a downstream element of FOXP3, allows the identification of LGALS3, a new marker of human regulatory T cells. Lab Invest 86, 724–737 (2006). https://doi.org/10.1038/labinvest.3700432
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/labinvest.3700432
Keywords
This article is cited by
-
Regulatory T cells in rheumatoid arthritis: functions, development, regulation, and therapeutic potential
Cellular and Molecular Life Sciences (2022)
-
CD98-induced CD147 signaling stabilizes the Foxp3 protein to maintain tissue homeostasis
Cellular & Molecular Immunology (2021)
-
The expression profile of the ubiquitin-like modifier FAT10 in immune cells suggests cell type-specific functions
Immunogenetics (2018)
-
Characterizing the glycome of the mammalian immune system
Immunology & Cell Biology (2008)
-
Natural Tregs, CD4+CD25+ inhibitory hybridomas, and their cell contact dependent suppression
Immunologic Research (2007)

{kind=link}
{kind=link}