
Abstract
Secreted morphogens induce distinct cellular responses in a concentration-dependent manner and act directly at a distance1,2,3,4,5,6,7. The existence of morphogens during mesoderm induction and patterning in vertebrates has been highly controversial, and it remains unknown whether endogenous mesoderm inducers act directly as morphogens8,9,10, function locally9 or act through relay mechanisms11,12. Here we test the morphogen properties of Cyclops and Squint—two Nodal-related transforming growth factor-β signals required for mesoderm formation and patterning in zebrafish13,14,15,16. Whereas different levels of both Squint and Cyclops can induce different downstream genes14,17,18,19, we find that only Squint can function directly at a distance. These results indicate that Squint acts as a secreted morphogen that does not require a relay mechanism.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Change history
01 August 2001
An Erratum to this paper has been published: https://doi.org/10.1038/35087633
References
Turing, A. M. The chemical basis of morphogenesis. Phil. Trans. R. Soc. Lond. B. 237, 37–72 (1952).
Wolpert, L. Positional information and the spatial pattern of cellular differentiation. J. Theor. Biol. 25, 1–47 (1969).
Nellen, D., Burke, R., Struhl, G. & Basler, K. Direct and long-range action of a DPP morphogen gradient. Cell 85, 357–368 (1996).
Lecuit, T. et al. Two distinct mechanisms for long-range patterning by Decapentaplegic in the Drosophila wing. Nature 381, 387–393 (1996).
Zecca, M., Basler, K. & Struhl, G. Direct and long-range action of a wingless morphogen gradient. Cell 87, 833–844 (1996).
Neumann, C. J. & Cohen, S. M. Long-range action of Wingless organizes the dorsal–ventral axis of the Drosophila wing. Development 124, 871–880 (1997).
Neumann, C. & Cohen, S. Morphogens and pattern formation. BioEssays 19, 721–729 (1997).
Gurdon, J. B., Harger, P., Mitchell A. & Lemaire, P. Activin signalling and response to a morphogen gradient. Nature 371, 487–492 (1994).
Jones, C. M., Armes, N. & Smith, J. C. Signalling by TGF-β family members: short-range effects of Xnr-2 and BMP-4 contrast with the long-range effects of activin. Curr. Biol. 6, 1468–1475 (1996).
McDowell, N., Zorn, A. M., Crease, D. J. & Gurdon, J. B. Activin has direct long-range signalling activity and can form a concentration gradient by diffusion. Curr. Biol. 7, 671–681 (1997).
Reilly, K. M. & Melton, D. A. Short-range signaling by candidate morphogens of the TGF beta family and evidence for a relay mechanism of induction. Cell 86, 743–754 (1996).
Rodaway, A. et al. Induction of the mesendoderm in the zebrafish germ ring by yolk cell-derived TGF-β family signals and discrimination of mesoderm and endoderm by FGF. Development 126, 3067–3078 (1999).
Feldman, B. et al. Zebrafish organizer development and germ-layer formation require nodal-related signals. Nature 395, 181–185 (1998).
Sampath, K. et al. Induction of the zebrafish ventral brain and floorplate requires cyclops/nodal signalling. Nature 395, 185–189 (1998).
Rebagliati, M. R., Toyama, R., Haffter, P. & Dawid, I. B. cyclops encodes a nodal-related factor involved in midline signaling. Proc. Natl Acad. Sci. USA 95, 9932–7 (1998).
Schier, A. F. & Shen, M. M. Nodal signalling in vertebrate development. Nature 403, 385–389 (2000).
Erter, C. E., Solnica-Krezel, L. & Wright, C. V. Zebrafish nodal-related 2 encodes an early mesendodermal inducer signaling from the extraembryonic yolk syncytial layer. Dev. Biol. 204, 361–372 (1998).
Gritsman, K., Talbot, W. S. & Schier, A. F. Nodal signaling patterns the organizer. Development 127, 921–932 (2000).
Thisse, B., Wright, C. V. & Thisse, C. Activin- and Nodal-related factors control antero-posterior patterning of the zebrafish embryo. Nature 403, 425–428 (2000).
Jones, C. M., Kuehn, M. R., Hogan, B. L., Smith, J. C. & Wright, C. V. Nodal-related signals induce axial mesoderm and dorsalize mesoderm during gastrulation. Development 121, 3651–3662 (1995).
Agius, E., Oelgeschlager, M., Wessely, O., Kemp, C. & De Robertis, E. M. Endodermal Nodal-related signals and mesoderm induction in Xenopus. Development 127, 1173–1183 (2000).
Gritsman, K. et al. The EGF-CFC protein one-eyed pinhead is essential for nodal signaling. Cell 97, 121–132 (1999).
Vogel, A. M. & Gerster, T. Promoter activity of the zebrafish bhikhari retroelement requires an intact activin signaling pathway. Mech. Dev. 85, 133–146 (1999).
Ruppert, R., Hoffmann, E. & Sebald, W. Human bone morphogenetic protein 2 contains a heparin-binding site which modifies its biological activity. Eur. J. Biochem. 237, 295–302 (1996).
Pfeiffer, S., Alexandre, C., Calleja, M. & Vincent, J. P. The progeny of wingless-expressing cells deliver the signal at a distance in Drosophila embryos. Curr. Biol. 10, 321–324 (2000).
Schier, A. F., Neuhauss, S. C., Helde, K, A., Talbot, W. S. & Driever, W. The one-eyed pinhead gene functions in mesoderm and endoderm formation in zebrafish and interacts with no tail. Development 124, 327–342 (1997).
Zhang, J., Talbot, W. S. & Schier, A. F. Positional cloning identifies zebrafish one-eyed pinhead as a permissive EGF-related ligand required during gastrulation. Cell 92, 241–251 (1998).
Strähle, U. et al. one-eyed pinhead is required for development of the ventral midline of the zebrafish (Danio rerio) neural tube. Genes Funct. 1, 131–148 (1997).
Acknowledgements
We thank members of the Schier and Yelon laboratories for discussions; R. Burdine, A. Carmany-Rampey, K. Joubin, R. Lehmann, G. Struhl, W. Talbot and D. Yelon for comments on the manuscript; S. Zimmerman, R. Feeney and T. Bruno for fish care. Y.C. is the Rebecca Ridley Kry Fellow of the Cancer Research Fund of the Damon Runyon–Walter Winchell Foundation. A.F.S. is a Scholar of the McKnight Endowment Fund for Neuroscience and the Irma T. Hirschl Trust, and is supported by grants from the NIH.
Author information
Authors and Affiliations
Corresponding author
Supplementary information

Supplementary Fig.1 (JPG 34 KB)
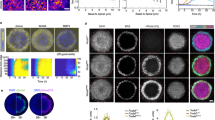
The long-range effect of Sqt is not a result of cell movement. a, illustration of the experiment in b-g; wild-type donor cells labeled with biotin-dextran and rhodamin-dextran (red) are transplanted at a distance from the sqt injected cells (green) at the 1000-2000 cell stage. b-d, b was taken shortly after transplantation; n and o were taken at 1hr (c) and 2hrs (d) after b; the three subsets of the transplanted cells are indicated by arrowheads of different colors. e-g, the embryo in e was fixed immediately after photographing and analyzed by in situ hybridization for ntl expression; red staining marks the transplanted cells produced by detecting the lineage tracer biotin-dextran with the ABC kit from Vectastain; the arrowheads in e-g correspond to those in b-d; three transplanted cells expressing ntl are indicated by the white arrows in g.
Rights and permissions
About this article
Cite this article
Chen, Y., Schier, A. The zebrafish Nodal signal Squint functions as a morphogen. Nature 411, 607–610 (2001). https://doi.org/10.1038/35079121
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/35079121
This article is cited by
-
Nodal is a short-range morphogen with activity that spreads through a relay mechanism in human gastruloids
Nature Communications (2022)
-
Single-molecule tracking of Nodal and Lefty in live zebrafish embryos supports hindered diffusion model
Nature Communications (2022)
-
Generation of extracellular morphogen gradients: the case for diffusion
Nature Reviews Genetics (2021)
-
MiR-430 Can Affect the Mesoderm Formation and Metamorphosis of Paralichthys olivaceus by Targeting lefty Gene
Journal of Ocean University of China (2020)
-
Studying molecular interactions in the intact organism: fluorescence correlation spectroscopy in the living zebrafish embryo
Histochemistry and Cell Biology (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}