
Abstract
Stomata are microscopic pores on the surfaces of leaves, the number and density of which vary in response to changes in environmental conditions, such as carbon dioxide concentration and light. We show here that mature leaves of Arabidopsis thaliana detect and transmit this external information to new leaves of the same plant, producing an appropriate adjustment of stomatal development. As CO2 concentration controls both stomatal opening1 and number2,3, and stomatal numbers also increase with higher light intensity4, the large gradients of CO2 and light found within plant communities5 have the potential to influence stomatal development.
Similar content being viewed by others

Main
Every year, 40% of the CO2 in the atmosphere passes through stomata6. Stomatal numbers modify both photosynthesis and efficient use of water1, and so any change in stomatal numbers7 in response to CO2 and light can influence photosynthesis and atmospheric CO2 concentration. The response mechanism described here may exert global effects that are not currently included in canopy and vegetation models of increasing atmospheric CO2 levels.
A genetic component in the response of stomatal development to increasing atmospheric CO2 concentration has been identified8 and the control of stomatal development by CO2 concentration is known to occur during early leaf development9,10, when ambient CO2 concentrations may not be accurately detectable by a new leaf sheathed by antecedent leaf primordia11. We therefore tested whether CO2 concentration can be detected by mature leaves in open ambient conditions, which might then transmit a signal to induce an appropriate developmental response by the stomata of new leaves. Our experimental design using the model plant Arabidopsis thaliana is shown in Fig. 1a.

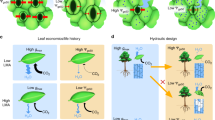
a, Leaf-cuvette experiment. Plants of Arabidopsis (Columbia, Col-0) were grown for 4 weeks under ambient CO2 (360 p.p.m.) until leaf insertions 5 to 13 had developed. These mature leaves were enclosed in transparent airtight cuvettes under CO2 concentrations of either 720 or 360 p.p.m. Subsequent leaf insertions developed outside the cuvette under ambient CO2. Plants were maintained in cuvettes for 7 to 9 days until the next five leaf insertions had matured, the last three of which were investigated for stomatal density (no. of stomata per mm2) and index ((no. of stomata/no. of stomata + no.of epidermal cells) × 100). b, (left) Stomatal index and density for new leaves (insertions 16 to 19) under ambient CO2 when mature leaves (insertions 5 to 13) inside cuvettes are supplied with increased CO2 (720 p.p.m.). Both stomatal density and index are reduced in new leaves if the supply of CO2 is increased to the mature leaves. Right, reverse experiment: mature leaves inside cuvettes are under CO2 at 360 p.p.m.; external CO2 is 720 p.p.m. Stomatal density and index increase in response to the decreased CO2 around the mature leaves. c, Effect on stomatal index of new leaves of reducing light incident on mature leaves by using neutral density filters (shade) or transparent filters (full light). Stomatal index of new leaves is reduced when mature leaves are shaded. *** P < 0.0005; * P < 0.05; bars, s.e.m.; n = 150.
Expanding leaves outside the cuvette (ambient CO2, 360 p.p.m.), with mature leaves exposed to a high concentration of CO2 inside the cuvette (720 p.p.m.; Fig. 1a), developed with a reduced stomatal index and density (Fig. 1b, left) compared with control plants grown entirely at ambient CO2. Reversing the cuvette arrangement so that the mature leaves were exposed to 360 p.p.m. and the expanding leaves to 720 p.p.m. CO2 resulted in a complete reversal (increase) of the stomatal index and density of the new leaves (Fig. 1b, right).
In these experiments, both abaxial (upper) and adaxial (lower) leaf surfaces responded in a similar manner, indicating that CO2 concentration is detected by mature leaves which signal to expanding leaves to induce an appropriate developmental response. To our knowledge, this is the first demonstration that mature leaves both detect CO2 concentration and transmit a long-distance signal that controls stomatal development in young leaves. Expanding leaves appeared to have no capacity to detect ambient CO2 concentration or to respond to it directly by altering stomatal initiation; no cuvette effect was found to contribute to the responses.
This mechanism of CO2 detection and signalling could enhance or optimize performance in plant communities. For example, the reduction in stomatal index and density with CO2 enrichment enhances water-use efficiency3,12, but such a response will be much less effective13 when leaves develop in the shade of other plants. We therefore tested whether the mechanisms controlling the response of stomatal initiation to CO2 might also be accompanied by another that responds to light, reducing the initiation of stomata with increasing shade.
We have previously shown that there is a positive relation in Arabidopsis between irradiance and both stomatal index and density. Fully grown leaves were now placed in shaded light, with the expanding leaves under full light. The stomatal index (Fig. 1c) and density (results not shown) of new leaves were reduced, as if they had developed under shade conditions. We conclude that long-distance signalling must also be involved in controlling the response of stomatal development to light, which supports the idea that there is an ecologically important link between the responses evoked by light and by carbon dioxide.
References
Berryman, C. A., Eamus, D. & Duff, G. A. J. Exp. Bot. 45, 539–546 (1994).
Woodward, F. I. Nature 327, 617–618 (1987).
Beerling, D. J., McElwain, J. C. & Osborne, C. P. J. Exp. Bot. 49, 1603–1607 (1998).
Tichà, I. Photosynthetica 11, 375–471 (1982).
Buchmann, N., Kao, W.-Y & Ehleringer, J. R. Glob. Change Biol. 2, 421–432 (1996).
Ciais, P. et al. J. Geophys. Res. Atmos. 102, 5857–5872 (1997).
Douville, H. et al. J. Geophys. Res. Atmos. 105, 14841–14861 (2000).
Gray, J. E. et al. Nature 408, 713–716 (2000).
Woodward, F. I. & Bazzaz, F. A. J. Exp. Bot. 39, 1771–1781 (1988).
Donnelly, P. M., Bonetta, D., Tsukaya, H., Dengler, R. E. & Dengler, N. G. Dev. Biol. 215, 407–419 (1999).
Jackson, D., Veit, B. & Hake, S. Development 120, 405–413 (1994).
Eamus, D., Berryman, C. A. & Duff, G. A. Aus. J. Pl. Physiol. 20, 741–755 (1993).
Muraoka, H., Tang, Y., Terashima, I., Koizumi, H. & Washitani, I. Plant Cell Environ. 23, 235–250 (2000).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lake, J., Quick, W., Beerling, D. et al. Signals from mature to new leaves. Nature 411, 154 (2001). https://doi.org/10.1038/35075660
Issue Date:
DOI: https://doi.org/10.1038/35075660
This article is cited by
-
Elevated CO2 Priming as a Sustainable Approach to Increasing Rice Tiller Number and Yield Potential
Rice (2023)
-
Integrating stomatal physiology and morphology: evolution of stomatal control and development of future crops
Oecologia (2021)
-
Dependency of Growth, Water Use Efficiency, Chlorophyll Fluorescence, and Stomatal Characteristics of Lettuce Plants to Light Intensity
Journal of Plant Growth Regulation (2021)
-
Hydrogen peroxide as a systemic messenger in the photosynthetic induction of mulberry leaves
Journal of Forestry Research (2021)
-
In vitro regeneration, antioxidant potential, and genetic fidelity analysis of Asystasia gangetica (L.) T.Anderson
In Vitro Cellular & Developmental Biology - Plant (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
