
Abstract
Asexual cells are normally able to reproduce entirely by themselves. But we have discovered that in about one-third of the dividing cells of Entamoeba invadens contraction of the cleavage furrow1 may stop before separation is complete. We show here that the connected daughter cells overcome this problem by calling upon a neighbouring amoeba to help them achieve the final stage of division. The 'midwife' cell is chemotactically recruited for this mechanical intervention in what is a surprising example of primitive cooperation.
Similar content being viewed by others

Main
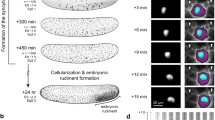
When an amoeba divides, the two daughter cells stay attached by a tubular tether which remains intact unless mechanically severed. If called upon, the neighbouring amoeba midwife (green cell in Fig. 1) travels up to 200 μm towards the dividing amoeba (see movie in supplementary information), usually advancing in a straight trajectory with an average velocity of 0.54 ± 0.08 μm s−1. The midwife then proceeds to rupture the connection, after which all three amoebae move on.

Frame times (a–i) are 0, 13, 30, 36, 41, 43, 49, 59 and 84 s, respectively. a–c, The midwife (green) aligns itself against the cleavage furrow; d–f, it puts out a pseudopod perpendicular to the furrow; g–i, then forcibly moves into the gap to disconnect the two daughter cells (blue). Although a few midwife amoebae may respond to the chemoattractant, only one, usually the nearest neighbour, is involved in completing the division. The midwife begins to move towards the dividing cell 2–3 min after division begins; this timing is compatible with an attractant with a diffusion coefficient of 10−6–10−7cm2 s−1 diffusing over a distance of 100 μm. Scale bar, 10 μm.
Using four different samples, each at amoeba densities of about 3×104 cells cm−2, we recorded 106 divisions, of which 32 (30±7%) were involved in cooperative reproduction (statistics were averaged over samples to give a 95% confidence level). In 11 cases (10−5%), the division was aborted, and the amoebae continued to live as multinucleated cells. The multinuclear cells may resume their attempt to divide, sometimes producing three or four viable daughter cells2,3.
In most cases (63 divisions, or 60±8%), however, separation eventually occurred4, with both daughter cells resuming motility and using traction force to move away from each other, tugging at their connection so that it lengthened and narrowed until it finally broke. The incidence of abortions and tripolar divisions rises significantly (20−5% aborted of 82 divisions) at low cell densities, when midwifing is rare.
To test whether the midwife is attracted chemotactically to the labouring division site, we aspirated fluid from the vicinity of the cleavage furrow of a dividing amoeba with a micropipette, then carefully discharged it near a distant amoeba. Of 41 such experiments, 15 (37%) revealed a positive chemotactic response (see movie in supplementary information; amoebae extended a directed pseudopod and followed a retracting pipette for a distance of over three times their own length); in 12 (30%), no response was seen; and in the remainder, cells moved in the general direction of the stimulus but resumed random motion before traversing 100 μm.
Amoebae showing positive chemotactic responses pursued the pipette over distances of a few hundred micrometres, persistently following the tip when we changed the direction of the pipette's trajectory. Controls consisting of fluid aspirated from the vicinity of a non-dividing amoeba or of fresh medium alone elicited no response from neighbouring cells.
Identifying the chemical attractant in a volume of only 10 picolitres is difficult, so we sampled a mature culture of cells at the peak of their division cycle, reasoning that the concentration of chemoattractant would be higher than in a young culture. We found that fluid drawn from a confluent culture (3–4 days old, undergoing over a million divisions per flask per day) and transferred into a culture of only a few hours old, induced a strong chemotactic response in the younger cells (see movie). These amoebae followed retreating pipettes for over 30 min at an average linear velocity of 0.73 ± 0.09 μm s−1 and flocked towards a stationary pipette at an average radial velocity of 0.34±0.04 μm s−1. Control solutions of cell-free three-day-old medium elicited no response; to control for any possible effect of starvation, fresh medium was changed 18 hours before collecting the fluid.
As we sampled fluid from cultures before the cells started clustering, we consider it most likely that the factor responsible for attracting midwives is the one secreted by cytokinesing cells, although it could be an entirely different chemoattractant. We have established that its relative molecular mass is between 50K and 100K and that it is still active after heating for 20 min at 95 °C, but it is sensitive to oxidation by 0.24 M metaperiodate. Glycoconjugates, particularly membrane lipophosphoglycan-like molecules5, have these properties and are therefore likely candidates for the midwife-recruiting chemoattractant molecule.
A similar phenomenon has recently been observed in dividing cells of the amoeba Dictyostelium discoideum (G. Gerisch and J. Köhler, personal communication). Midwifing may therefore constitute a more general adaptation of existing chemotactic mechanisms6,7, enabling flaws in reproduction to be overcome through cooperation.
References
Schroeder, T. E. Ann. NY Acad. Sci. 582, 78–87 (1990).
Weber, I. et al. Curr. Biol. 10, 501–506 (2000).
Makioka, A., Kumagai, M., Ohtomo, H., Kobayashi, S. & Takeuchi, T. Parasitol. Res. 86, 599–602 (2000).
Fukui, Y. Microscopy Res. Tech. 49, 183–189 (2000).
Moody-Haupt, S., Patterson, J. H., Mirelman, D. & McConville, M. J. J. Mol. Biol. 297, 409–420 (2000).
Bailey, G. B., Leitch, G. J. & Day, D. B. J. Protozool. 32, 341–346 (1985).
Mato, T. M., Losada, A., Nanjundiah, V. & Konijn, T. M. Proc. Natl Acad. Sci. USA 72, 4991–4993 (1975).
Author information
Authors and Affiliations
Corresponding author
Supplementary information
Midwife event imaged at 10X magnification.
At this magnification the full trajectory is visualized. Time between frames is 0.4 seconds. (MOV 922 kb)
Typical midwife event images at 100X magnification.
The original location from which the midwife approaches is not visible due to the large magnification. Note the abrupt and directed pseudopod extension into the cleavage furrow. Time between frames is 0.4 seconds. (MOV 1189 kb)
Chemotactic attraction towards a retracting pipette.
Fluid was first collected around a dividing cell, then gradually ejected at a different site. White arrow marks the amoeba with the strongest chemotactic response. Black arrow indicates distance traversed by pipette. Total time before editing was 6 minutes. (MOV 2018 kb)
Chemotactic attraction towards a stationary pipette.
Fluid was collected from a confluent culture, and ejected into a low-density freshly plated one. Within minutes aggregation of distant amoebae was evident. Time between frames is 15 seconds. (MOV 859 kb)
*All formats are Quicktime movies, produced by NIH Image v1.62 on a Macintosh G3 using "Greyscale", "Photo-JPEG" compression, and "Low" quality, run at 8 frames per second.
Rights and permissions
About this article
Cite this article
Biron, D., Libros, P., Sagi, D. et al. 'Midwives' assist dividing amoebae. Nature 410, 430 (2001). https://doi.org/10.1038/35068628
Issue Date:
DOI: https://doi.org/10.1038/35068628
This article is cited by
-
Behavior of DNA-lacking mitochondria in Entamoeba histolytica revealed by organelle transplant
Scientific Reports (2017)
-
A novel mode of cytokinesis without cell-substratum adhesion
Scientific Reports (2017)
-
Genome Re-duplication and Irregular Segregation Occur During the Cell Cycle of Entamoeba histolytica
Bioscience Reports (2007)
-
Midwifery and assisted reproduction inDictyostelium andEntamoeba
Journal of Biosciences (2003)
-
Eukaryotic checkpoints are absent in the cell division cycle ofEntamoeba histolytica
Journal of Biosciences (2002)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
