
Abstract
The congregation of locusts into vast swarms can cause crop devastation of biblical proportions1. Here we show that guaiacol, a key component of a pheromone derived from locust faecal pellets that promotes the aggregation of locusts2,3,4,5, is produced by bacteria in the locust gut. This adaptation by an insect to exploit a common metabolite produced by indigenous gut bacteria has wide implications for our appreciation of the role of the gut microbiota in insects.
Similar content being viewed by others

Main
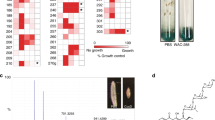
Guaiacol (2-methoxyphenol) and phenol are volatile compounds that are both released from the faecal pellets of conventionally reared larval and mature adult desert locusts, Schistocerca gregaria (Table 1). Guaiacol production from locusts has previously been attributed to the insect itself. However, desert locusts have a large but relatively simple gut bacterial biota6 which comprises bacterial species acquired from their environment and located in particular in the lining of the hindgut, where faecal pellets form.
We investigated the possible involvement of the gut biota in the production of guaiacol by rearing locusts from surface-sterilized eggs in a sterile isolator system and establishing a breeding colony of axenic (germ-free) locusts by feeding them γ-irradiated freeze-dried grass and bran7. Faecal pellets from axenic locusts smelled markedly different from those from locusts with a normal gut biota. Chemical analysis revealed that the difference in odour was due to the absence of guaiacol and low levels of phenol in volatiles released from the germ-free faecal pellets.
We detected guaiacol and phenol in volatiles from the faecal pellets of monoassociated fifth-instar larvae, immature and mature adults carrying a single bacterial species Pantoea (=Enterobacter) agglomerans, a prominent member of the locust-gut biota6, and reared on the γ-irradiated diet (Table 1). The smaller amount of volatile phenolics released from young-adult monoassociated insects in comparison with other stages correlates with the lower numbers of bacteria in the gut of newly moulted locusts6. These results show that guaiacol originates from the gut bacteria. This finding is supported by experiments demonstrating that three different species of bacteria (including P. agglomerans) from the locust gut can produce guaiacol directly from axenic faecal pellets in vitro.
The precursor for guaiacol synthesis in faecal pellets must either be a component of plant material or an excretory product of the insect itself. The former is most likely, as the amount of guaiacol produced depends on the diet: more guaiacol was produced by normal locusts fed on fresh wheat seedlings than by those fed freeze-dried γ-irradiated grass. Incubation of locust food with bacteria generated only trace amounts of guaiacol or phenol (data not shown), indicating that digestion of the plant material in the locust gut is required for production of guaiacol by the bacteria.
The most likely precursor for guaiacol synthesis is lignin-derived vanillic acid (4-hydroxy-3-methoxybenzoic acid), which is found in the faeces of both axenic and normal locusts8. Microbial transformation of vanillic acid to guaiacol requires a decarboxylation step9. Consistent with this, we found guaiacol released by all three species of bacteria from glucose/peptone broth cultures containing vanillic acid (data not shown). Furthermore, faeces from conventionally reared insects fed filter paper impregnated with vanillic acid solution yielded large amounts of guaiacol (Table 1).
The gut bacteria of locusts, as in many other insects, are considered to be either commensal or facultatively pathogenic10, and therefore to have little effect on their hosts. Our results show that locusts have adapted to use a pheromonal component that is derived from its digestive waste products by the action of bacteria acquired serendipitously with its food. The gut bacteria also help the locust to defend itself against microbial pathogens, mainly by producing antimicrobial phenolic compounds8,11,12,13. These contributions by the insect's gut microbiota to its behaviour and survival were previously unsuspected.
References
Steedman, A. Locust Handbook 2nd edn 2–9 (Overseas Development Natural Resources Institute, London, 1988).
Fuzeau-Braesch, S., Genin, E., Jullien, R., Knowles, E. & Papin, C. J. Chem. Ecol. 14, 1023–1033 (1988).
Obeng-Ofori, D., Torto, B., Njagi, P. G. N., Hassanali, A. & Amiani, H. J. Chem. Ecol. 20, 2077–2087 (1994).
Byers, J. A. Biol. Rev. 66, 347–378 (1991).
Pener, M. P. & Yerushalmi, Y. J. Insect Physiol. 44, 365–377 (1998).
Hunt, J. & Charnley, A. K. J. Invertebr. Pathol. 38, 378–385 (1981).
Charnley, A. K., Hunt, J. & Dillon, R. J. J. Insect Physiol. 31, 477–485 (1985).
Dillon, R. J. & Charnley, A. K. Can. J. Microbiol. 34, 1075–1082 (1988).
Crawford, R. L. & Olson, P. P. Appl. Environ. Microbiol. 36, 539–543 (1978).
Lysenko, O. Annu. Rev. Microbiol. 39, 673–695 (1985).
Dillon, R. J. & Charnley, A. K. J. Invertebr. Pathol. 47, 350–360 (1986).
Dillon, R. J. & Charnley, A. K. in The Fungal Spore and Disease Initiation in Plants and Animals (eds Cole, G. T. & Hoch, H. C.) 129–156 (Plenum, New York, 1991).
Dillon, R. J. & Charnley, A. K. J. Invertebr. Pathol. 66, 72–75 (1995).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dillon, R., Vennard, C. & Charnley, A. Exploitation of gut bacteria in the locust. Nature 403, 851 (2000). https://doi.org/10.1038/35002669
Issue Date:
DOI: https://doi.org/10.1038/35002669
This article is cited by
-
Effect of antibiotic treatment and gamma-irradiation on cuticular hydrocarbon profiles and mate choice in tsetse flies (Glossina m. morsitans)
BMC Microbiology (2018)
-
Pyrazines from bacteria and ants: convergent chemistry within an ecological niche
Scientific Reports (2018)
-
Stingless Bee Larvae Require Fungal Steroid to Pupate
Scientific Reports (2018)
-
Microbiota in insect fungal pathology
Applied Microbiology and Biotechnology (2018)
-
The evolutionary and coevolutionary consequences of defensive microbes for host-parasite interactions
BMC Evolutionary Biology (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
