
Abstract
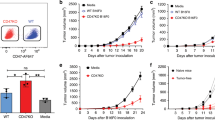
The interaction between CD40 ligand (CD40L, CD154) and its receptor CD40 on antigen-presenting cells, is essential for the initiation of cell-mediated and humoral immune responses. In this study, we investigated the antitumor effect of in vivo gene transfer of CD40L to tumor cells using an adenoviral vector (AdCMVmCD40L) in a murine CT-26 colon cancer model. We found that injection of AdCMVmCD40L caused tumor regression in a dose-dependent manner. A complete regression of tumor was observed in 81% of mice treated with 109 p.f.u. of AdCMVmCD40L. The antitumor effect induced by CD40L was mediated by CD8+ T cells and was associated with the generation of tumor-specific cytolytic T lymphocytes (CTL). Animals that eradicated the tumor were protected against tumor cell rechallenge, and both CD4+ and CD8+ T cells were involved in specific protective immunity. Treatment with AdCMVmCD40L in one tumor nodule also caused complete regression of established tumors at distant sites. The antitumor effect elicited by AdCMVmCD40L was associated with the intratumoral production of IL-12 and IFN-γ and with an increased intratumoral expression of chemokines such as MIP-1α, MIP-1β, MIP-2, RANTES, and eotaxin. These data demonstrate that intratumoral injection of AdCMVmCD40L induces a powerful cascade of chemokines and cytokines in the tumor mass and stimulates an efficient antitumor immunity leading to regression of established colon cancer and protection against tumor cell rechallenge.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout










Similar content being viewed by others


References
Noelle RJ, Ledbetter JA, Aruffo A . CD40 and its ligand, an essential ligand–receptor pair for thymus-dependent B cell activation Immunol Today 1992 13: 431–433
Grewal IS, Flavell RA . CD40 and CD154 in cell-mediated immunity Annu Rev Immunol 1998 16: 111–135
Roy M et al. The regulation of the expression of gp39, the CD40 ligand, on normal and cloned CD4+ T cells J Immunol 1993 151: 2497–2510
Gray D, Siepmann K, Wohlleben G . CD40 ligation in B cell activation, isotype switching and memory development Semin Immunol 1994 6: 303–310
Hollenbaugh D et al. The role of CD40 and its ligand in the regulation of the immune response Immunol Rev 1994 138: 23–37
Ridge JP, Rosa FD, Matzinger P . A conditioned dendritic cell can be a temporal bridge between a CD4+ T-helper and a T-killer cell Nature 1998 393: 474–478
Bennett SR et al. Help for cytotoxic T cell responses is mediated by CD40 signalling Nature 1998 393: 478–480
Schoenberger SP et al. T cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions Nature 1998 393: 480–483
Caux C et al. Activation of human dendritic cells through CD40 cross-linking J Exp Med 1994 180: 1263–1272
Peguet-Navarro J et al. Functional expression of CD40 antigen on human epidermal Langerhans cells J Immunol 1995 155: 4241–4247
Cella M et al. Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T–T help via APC activation J Exp Med 1996 184: 747–752
Koch F et al. High level IL-12 production by murine dendritic cells: up-regulation via MHC class II and CD40 molecules and downregulation by IL-10 J Exp Med 1996 184: 741–746
Ludewig B et al. Spontaneous apoptosis of dendritic cells is efficiently inhibited by TRAP (CD40-ligand) and TNF-alpha, but strongly enhanced by interleukin-10 Eur J Immunol 1995 25: 1943–1950
Costello RT, Gastaut JA, Olive D . What is the real role of CD40 in cancer immunotherapy Immunol Today 1999 20: 488–493
Wang D et al. Role of the CD40 and CD95 (APO-1/Fas) antigens in the apoptosis of human B cell malignancies Br J Haematol 1997 97: 409–417
Fluckiger AC, Durand I, Banchereau J . Interleukin 10 induces apoptotic cell death of B-chronic lymphocytic leukemia cells J Exp Med 1994 179: 91–99
Hirano A et al. Inhibition of human breast carcinoma growth by a soluble recombinant human CD40 ligand Blood 1999 93: 2999–3007
Sotomayor EM et al. Conversion of tumor-specific CD4+ T cell tolerance to T cell priming through in vivo ligation of CD40 Nature Med 1999 5: 780–787
Diehl L et al. CD40 activation in vivo overcomes peptide-induced peripheral cytotoxic T-lymphocyte tolerance and augments anti-tumor vaccine efficacy Nature Med 1999 5: 774–779
French RR, Chan HT, Tutt AL, Glennie MJ . CD40 antibody evokes a cytotoxic T cell response that eradicates lymphoma and bypasses T cell help Nature Med 1999 5: 548–553
Dilloo D et al. CD40 ligand induces an antileukemia immune response in vivo Blood 1997 90: 1927–1933
Grossmann ME, Brown MP, Bernner MK . Antitumor responses induced by transgenic expression of CD40 ligand Hum Gene Ther 1997 8: 1935–1943
Couderc B et al. Enhancement of antitumor immunity by expression of CD70 (CD27 ligand) or CD154 (CD40 ligand) costimulatory molecules in tumor cells Cancer Gene Ther 1998 5: 163–175
Nakajima A et al. Antitumor effect of CD40 ligand: elicitation of local and systemic antitumor responses by IL-12 and B7 J Immunol 1998 161: 1901–1907
Kato K, Cantwell MJ, Sharma S, Kipps TJ . Gene transfer of CD40-ligand induces autologous immune recognition of chronic lymphocytic leukemia B cells J Clin Invest 1998 101: 1133–1141
Kikuchi T, Crystal RG . Anti-tumor immunity induced by in vivo adenovirus vector-mediated expression of CD40 ligand in tumor cells Hum Gene Ther 1999 10: 1375–1387
Trinchieri G . Interleukin-12 and its role in the generation of TH1 cells Immunol Today 1993 14: 335–338
Nastala CL et al. Recombinant IL-12 administration induces tumor regression in association with IFN-gamma production J Immunol 1994 153: 1697–1706
Chiodoni C et al. Dendritic cells infiltrating tumors cotransduced with granulocyte/macrophage colony-stimulating factor (GM-CSF) and CD40 ligand genes take up and present endogenous tumor-associated antigens, and prime naive mice for a cytotoxic T lymphocyte response J Exp Med 1999 190: 125–133
Gurunathan S et al. CD40 ligand/trimer DNA enhances both humoral and cellular immune responses and induces protective immunity to infectious and tumor challenge J Immunol 1998 161: 4563–4571
Esche C et al. CD154 inhibits tumor-induced apoptosis in dendritic cells and tumor growth Eur J Immunol 1999 29: 2148–2155
Hermans IF et al. Impaired ability of MHC class II−/− dendritic cells to provide tumor protection is rescued by CD40 ligation J Immunol 1999 163: 77–81
Mackey MF et al. Cutting edge: dendritic cells require maturation via CD40 to generate protective antiumor immunity J Immunol 1998 161: 2094–2098
Mazzolini G et al. Expression of colon cancer and induction of antitumor immunity by intratumoral injection of adenovirus expressing inteleukin-12 Cancer Gene Ther 1999 6: 514–522
Baggiolini M . Chemokines and leukocyte traffic Nature 1998 392: 565–568
Mantovani A . The chemokine system: redundancy for robust outputs Immunol Today 1999 20: 254–257
Dieu MC et al. Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites J Exp Med 1998 188: 373–386
Taub DD et al. Preferential migration of activated CD4+ and CD8+ T cells in response to MIP-1α and MIP-1β Science 1993 260: 355–358
Zou LP et al. Dynamics of production of MIP-1alpha, MCP-1 and MIP-2 and potential role of neutralization of these chemokines in the regulation of immune responses during experimental autoimmune neuritis in Lewis rats J Neuroimmunol 1999 98: 168–175
Diab A et al. Neutralization of macrophage inflammatory protein 2 (MIP-2) and MIP-1α attenuates neutrophil recruitment in the central nervous system during experimental bacterial meningitis Infect Immun 1999 67: 2590–2601
Hang L et al. Macrophage inflammatory protein-2 is required for neutrophil passage across the epithelial barrier of the infected urinary tract J Immunol 1999 162: 3037–3044
Bergelson JM et al. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5 Science 1997 275: 1320–1323
Wickham TJ, Mathias P, Cheresh DA, Nemerow GR . Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment Cell 1993 73: 309–319
Brattain MG et al. Establishment of mouse colonic carcinoma cell lines with different metastatic properties Cancer Res 1980 40: 2142–2146
Qian C, Bilbao R, Bruña O, Prieto J . Induction of sensitivity to ganciclovir in human hepatocellular carcinoma cells by adenovirus-mediated gene transfer of herpes simplex virus thymidine kinase Hepatology 1995 22: 118–123
Lasarte JJ et al. Different doses of adenoviral vector expressing IL-12 enhance or depress the immune response to a coadministered antigen: the role of nitric oxide J Immunol 1999 162: 5270–5277
Acknowledgements
This work was supported in part by SAF 98-0146 from CICYT, Inés Bemberg Grant and also by J Vidal, Dr Cervera and M Mendez grants for Gene Therapy.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Sun, Y., Peng, D., Lecanda, J. et al. In vivo gene transfer of CD40 ligand into colon cancer cells induces local production of cytokines and chemokines, tumor eradication and protective antitumor immunity. Gene Ther 7, 1467–1476 (2000). https://doi.org/10.1038/sj.gt.3301264
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.gt.3301264
Keywords
This article is cited by
-
Enhanced therapeutic anti-tumor immunity induced by co-administration of 5-fluorouracil and adenovirus expressing CD40 ligand
Cancer Immunology, Immunotherapy (2014)
-
Immunotherapy for liver tumors: present status and future prospects
Journal of Biomedical Science (2009)
-
Adenovirus delivery of human CD40 ligand gene confers direct therapeutic effects on carcinomas
Cancer Gene Therapy (2009)
-
A Tumor-selective Biotherapy With Prolonged Impact on Established Metastases Based on Cytokine Gene-engineered MSCs
Molecular Therapy (2008)
-
A Tumor-selective Biotherapy With Prolonged Impact on Established Metastases Based on Cytokine Gene-engineered MSCs
Molecular Therapy (2008)
