
Abstract
Tolerance is defined as a decrease in responsiveness to a drug after repeated administration. Tolerance to the behavioral effects of hallucinogens occurs in humans and animals. In this study, we used drug discrimination to establish a behavioral model of lysergic acid diethylamide (LSD) tolerance and examined whether tolerance to the stimulus properties of LSD is related to altered serotonin receptor signaling. Rats were trained to discriminate 60 μg/kg LSD from saline in a two-lever drug discrimination paradigm. Two groups of animals were assigned to either chronic saline treatment or chronic LSD treatment. For chronic treatment, rats from each group were injected once per day with either 130 μg/kg LSD or saline for 5 days. Rats were tested for their ability to discriminate either saline or 60 μg/kg LSD, 24 h after the last chronic injection. Rats receiving chronic LSD showed a 44% reduction in LSD lever selection, while rats receiving chronic vehicle showed no change in percent choice on the LSD lever. In another group of rats receiving the identical chronic LSD treatment, LSD-stimulated [35S]GTPγS binding, an index of G-protein coupling, was measured in the rat brain by autoradiography. After chronic LSD, a significant reduction in LSD-stimulated [35S]GTPγS binding was observed in the medial prefrontal cortex and anterior cingulate cortex. Furthermore, chronic LSD produced a significant reduction in 2,5-dimethoxy-4-iodoamphetamine-stimulated [35S]GTPγS binding in medial prefrontal cortex and anterior cingulate cortex, which was blocked by MDL 100907, a selective 5-HT2A receptor antagonist, but not SB206553, a 5-HT2C receptor antagonist, indicating a reduction in 5-HT2A receptor signaling. 125I-LSD binding to 5-HT2A receptors was reduced in cortical regions, demonstrating a reduction in 5-HT2A receptor density. Taken together, these results indicate that adaptive changes in LSD-stimulated serotonin receptor signaling may mediate tolerance to the discriminative stimulus effects of LSD.
Similar content being viewed by others


INTRODUCTION
The hallucinogen, lysergic acid diethylamide (LSD), is a potent drug that produces altered perceptions, behavior, and mood in humans. In general, repeated drug use can lead to many psychological and physical consequences such as dependence and tolerance. Tolerance is often defined as a decrease in responsiveness to a drug after repeated administration. The underlying neuronal mechanism of tolerance is poorly understood. Tolerance develops to some of the behavioral effects of hallucinogens in animals. For example, tolerance develops to the disruptive effects of LSD on climbing behavior and fixed ratio lever pressing, and LSD-induced neophobia (Freedman et al, 1958, 1964; Winter, 1971; Adams and Geyer, 1985). In addition, the substituted amphetamine hallucinogens also produce tolerance. For example, repeated (1-(2,5-dimethoxy-4-methylphenyl)-2-aminopropane (DOM) treatments result in a decrease in DOM-induced head twitches in rats (Leysen et al, 1989). Finally, in animals trained to discriminate 2,5-dimethoxy-4-iodoamphetamine (DOI) from saline, our laboratory has demonstrated a reduction in percent DOI lever responding in rats chronically treated with DOI, while no changes were found in rats receiving chronic saline (Smith et al, 1999).
LSD displays high affinity for serotonin-2A (5-HT2A) and serotonin-2C (5-HT2C) receptors (Titeler et al, 1988). Although LSD interacts with other receptors (the 5-HT1 family, 5-HT5, 5HT6, 5HT7, α-adrenergic, and dopaminergic receptors), activation of 5-HT2A receptors is thought to mediate the hallucinogenic properties of LSD. In animals, the 5-HT2A receptor is considered an important mediator of the effects of hallucinogens based on behavioral studies. For example, 5-HT2A receptor agonists substitute for and selective 5-HT2A receptor antagonists block the LSD-induced discriminative stimulus (Glennon et al, 1984; Colpaert et al, 1985; Glennon and Hauck, 1985; Cunningham and Appel, 1987; Callahan and Appel, 1988; Fiorella et al, 1995; Smith et al, 1995; Appel et al, 2004). In addition, there is a significant correlation between the binding of agonists at the 5-HT2A receptor and potency in drug discrimination (Glennon et al, 1984). Finally, human studies report that most of the hallucinogenic properties of psilocybin, an indoleamine hallucinogen, are mediated by 5-HT2A receptors (Vollenweider et al, 1997, 1998). Thus, 5-HT2A receptor activation is proposed to mediate the behavioral and subjective effects of LSD and other hallucinogens.
In general, decreases in 5-HT2A receptor density have been reported after repeated administration of hallucinogens. For example, Leysen et al (1989) reported that repeated DOM produced significant decreases in 5-HT2A receptor density in the rat frontal cortex. Furthermore, Buckholtz et al (1990) demonstrated that after 5 days of LSD treatment, 3H-ketanserin binding is reduced in the cortex, hippocampus, midbrain, and brainstem, indicating decreased 5-HT2A and/or 5-HT2C receptor densities. Interestingly, other receptor subtypes were not altered such as 5-HT1A, 5-HT1B, α-adrenergic, β-adrenergic, D2 dopaminergic, further supporting the role of 5-HT2A receptors in the actions of LSD (Buckholtz et al, 1990). Moreover, our laboratory demonstrated, using the same drug dosing regimen that produced tolerance to the stimulus effects of DOI, significant reductions in the density of brain 5-HT2A receptors in the cortex and claustrum (Cl) but not 5-HT2C receptors in the choriod plexus or striatum (STR) (Smith et al, 1999). These studies provide converging evidence that 5-HT2A receptors may be important in the stimulus properties of hallucinogens and suggest that adaptive changes in 5-HT2A receptor number may mediate tolerance to hallucinogens.
The goals of the present study were to determine (1) if tolerance develops to the stimulus properties of LSD and (2) if changes in serotonin receptor signaling occur that correspond to the development of tolerance. Radioligand binding experiments have several limitations. Binding sites might not represent functional receptors due to receptor reserve or intracellular sequestration. In addition, changes in receptor binding may be offset by changes in receptor function at the level of receptor–G-protein interaction. For this reason, we utilized [35S]GTPγS autoradiography to quantify functional receptor–G-protein interactions in brain sections after chronic LSD exposure. Receptor-stimulated [35S]GTPγS turnover, which measures the exchange of GDP for [35S]GTPγS, is a direct assay of receptor activation of G-proteins (Sim et al, 1997a). Since GTPγS is metabolically stable, [35S]GTPγS autoradiography allows the measurement of receptor–G-protein interaction with neuroanatomical resolution (Sim et al, 1997a). This technique has been shown to measure agonist-stimulated 5-HT2A receptor–Gαq coupling (Adlersberg et al, 2000). The present study demonstrates LSD-induced behavioral tolerance in drug discrimination and explores the functional state of 5-HT2A receptors to investigate the mechanisms mediating tolerance to LSD.
MATERIALS AND METHODS
Animals
Adult male Sprague–Dawley rats (225–249 g; Harlan Sprague–Dawley Inc., Indianapolis, IN) were individually housed and food deprived to 85% of their free-feeding weight 1 week prior to the beginning of the drug discrimination experiments. All animals had access to water except during training, and were given enough food immediately following training and on weekends to maintain their weights at 85% of their expected nondeprived weights. Rats were maintained in a colony room (ambient temperature 22–23°C, 12 : 12 light : dark cycle). For autoradiography studies, adult male Sprague–Dawley rats (225–249 g) were used and chronic injections were initiated 1 week after arrival. All animals were maintained in a colony room throughout the injection period with food and water available ad libitum. All animal use procedures were in strict accordance with the NIH Guide to the Care and Use of Laboratory Animals and approved by Vanderbilt University Animal Care Committee.
Apparatus
Six commercially available operant conditioning chambers (BRS/LVE Model RTC-024), each housed in a sound-attenuated chamber, were used. The operant chambers were equipped with two response levers, a liquid dipper centered between the two levers, and a house light. The equipment and experimental parameters were programmed using MED Associates software and controlled by MED Associates interface and MS-DOS-compatible computers.
Drug Discrimination and Behavioral Tolerance Studies
Rats (n=60) were shaped to lever press for milk reinforcement (Borden's Condensed Milk diluted 1 : 1 with tap water) on a Fixed Ratio 1 schedule during daily 20 min training sessions. After shaping to lever press, the reinforcement contingency was changed to a variable interval (VI) 15 s schedule of reinforcement with a 15 s timeout (TO). The TO contingency, a 15 s period after incorrect responses during which no reinforcement was available, served to punish incorrect responses. At this point, discrimination training began. The VI schedule was changed to a VI–30 s schedule at the end of the first week of training where it remained for the duration of the experiment. Rats were given 20 min training sessions Monday–Friday. At 30 min following injection with either 60 μg/kg LSD or saline, the animal was placed in the operant chamber and reinforced for responding on the correct lever. For half the animals, responding on the right lever was LSD correct and responding on the left lever was saline correct; the reverse was true for the remainder of the animals. Throughout training, LSD and saline were altered every other day. Discrimination learning was monitored twice a week by calculating the percent correct lever responses (number of responses on the correct lever/total number of responses) during 2.5 min test periods given at the beginning of the training session. Training continued until choice behavior for the whole group averaged 85% correct or greater for both LSD and saline during the 2.5 min extinction tests.
Following acquisition of the LSD–saline discrimination, a dose–response curve was determined during 5 min extinction test sessions. For this experiment, 40 rats were assigned to one of four groups (n=10). Rats were injected with LSD (15, 30, or 60 μg/kg) or saline 30 min prior to testing.
Following the dose–response experiment and a week of retraining, a study was performed to determine if tolerance would develop to the discriminative stimulus effects of LSD. The animals were assigned to one of four groups (n=12) and treated chronically with either LSD or vehicle (saline). Prior to chronic treatment, two groups of rats were tested for choice behavior following vehicle administration while the remaining two were tested following 60 μg/kg LSD. The preinjection interval was 30 min and all animals were tested during 5-min extinction sessions. One group of animals from each pretest condition (LSD–vehicle) was then assigned to chronic vehicle or chronic LSD treatment group. The rats were injected once per day with 130 μg/kg LSD s.c. or vehicle for 5 days. At 24 h following the last chronic injection, the animals were retested following either vehicle or 60 μg/kg LSD during 5-min extinction test sessions. No training was given during the 5-day treatment period.
Autoradiography Studies: Chronic Drug Treatment
Rats were injected daily (0900) with 130 μg/kg s.c. LSD or vehicle for 5 days and killed 24 h after the last dose.
Tissue Preparation
Rats were decapitated; brains were quickly removed, frozen in cold isopentane (−25°C), and stored at −80°C until sectioning. Frozen brains were sectioned at 20 μm thickness on a cryostat, thaw mounted on poly-L-lysine-subbed slides, dried under vacuum, desiccated, and stored at −80°C.
[35S]GTPγS Autoradiography
Under vacuum, sections were brought to room temperature and desiccated, and then incubated in assay buffer (50 mM Tris/HCl, 5 mM MgCl2, 0.2 mM EGTA, 100 mM NaCl, 0.2 mM DTT, 5 μg/ml leupeptin, 1 μg/ml pepstatin, 5 μg/ml aprotinin, 0.1 mM PMSF, pH 7.4) for 30 min at room temperature, followed by incubation with 2 mM GDP in assay buffer for 20 min at room temperature. 8-Cyclopentyl-1,3-dipropylxanthine, an adenosine A1 receptor antagonist (1 μM), was also added to the assay buffer to block the basal adenosine A1 receptor activity (Laitinen, 1999). For agonist stimulation, sections were incubated with agonist (5 nM LSD or 5 nM DOI), 0.04–0.05 nM [35S]GTPγS, and 2 mM GDP for 2 h at room temperature. For antagonist studies, sections were preincubated for 10 min in the presence of 10 nM MDL 100907, a selective 5-HT2A receptor antagonist, or 10 nM SB206553, a 5-HT2C receptor antagonist, prior to addition of 5 nM DOI. Basal activity was determined in the absence of agonist, and nonspecific binding was determined in the presence of unlabeled GTPγS (10 μM). Termination of incubation was accomplished by 3 × 3 min ice-cold washes with 50 mM Tris/HCl buffer pH 7.4, followed by rinsing with cold Milli-Q H2O. Air-dried sections were placed against Biomax film with 14C standard strip for 2–5 days.
125I-LSD Autoradiography
Brain sections were preincubated for 15 min at room temperature in assay buffer (170 mM Tris/HCl buffer, pH 7.4) containing 1 μM sulpiride and 1 μM SB206553 to mask dopamine D2 receptors and 5-HT2C receptors, respectively. Next, sections were incubated with assay buffer plus 0.5 nM 125I-LSD for 60 min at room temperature. Nonspecific binding was defined by incubating adjacent sections in the presence of 30 μM methysergide. Incubation was terminated by 3 × 3 min ice-cold washes with assay buffer followed by rinsing with cold Milli-Q H2O. Air-dried sections were placed against Biomax film with 14C standard strip for 1–3 days. To evaluate whether residual LSD after chronic LSD administration contributes to the decreased binding, we examined 125I-LSD binding at several different concentrations using mounted brain sections from chronic LSD- and saline-treated animals. There was a statistically significant reduction when high concentrations of 125I-LSD were used (⩾0.5 nM, presumably at or near BMAX for 125I-LSD binding) but not at a low concentration (eg, 0.125 nM). We interpret this as supporting the assertion that the decrease reported in the various tables after chronic LSD reflects a reduction in the density of 5-HT2A receptor binding sites.
Image Analysis
Autoradiograms were digitized using an AGFA duoscan T1200 scanner and Fotolook 3.6 software. Brain regions that were examined are illustrated in Figure 1. Regional analysis of [35S]GTPγS binding signal was performed using NIH Image J version 1.326 (Wayne Rasband, NIH).

Schematic representations of brain regions analyzed. Outlined areas highlight regions of quantification; see Materials and methods for details. Abbreviations used: ACC, anterior cingulate cortex; Cl, claustrum; En, endopiriform nucleus; FPC-M, frontal parietal cortex-motor; FPC-S, frontal parietal cortex-somatosensory; mPFC, medial prefrontal cortex; nA-C, nucleus accumbens-core; nA-S, nucleus accumbens-shell; PFC, prefrontal cortex; S, septum; STR, striatum; all measurements are relative to bregma (Paxinos and Watson, 1986).
Statistical Analysis
The behavioral data were analyzed using a two-way ANOVA. Level of significance for the ANOVA was set at P<0.05. Contribution of individual group means to the overall significant F-value was determined by Student–Newman–Keuls post hoc test (P<0.05). Autoradiographic data comparisons were made by one-way ANOVA. Level of significance for the ANOVA was set at P<0.05. Contribution of individual group means to the overall significant F-value was determined by Fisher LSD post hoc test (P<0.05). Statistical analyses were preformed with SPSS 7.0 software.
RESULTS
Effect of Chronic LSD Treatment on Choice Behavior
After 54 training sessions on the LSD (60 μg/kg) vs saline discrimination, the animals were averaging 85% correct or greater during the weekly 2.5 min extinction test sessions. The data from the 2.5 min extinction sessions averaged across the final four extinction periods (two LSD and two saline) were 92% correct for LSD and 85% correct for saline. A dose–response curve was determined for several doses of LSD and saline. LSD lever responding was orderly and dose-dependent; a dose of 29 μg/mg gave 50% responding.
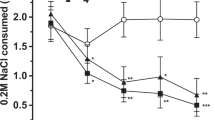
The effects of chronic LSD or saline on LSD discrimination are shown in Figure 2. The groups are identified by the chronic treatment condition (LSD or vehicle) and the test condition: for example, rats treated chronically with LSD and tested on LSD are labeled LSD-LSD, while rats treated chronically with vehicle and tested on LSD are labeled VEH-LSD. Prior to chronic treatment (pre-tx), neither the two groups of rats tested on vehicle nor the two groups tested on LSD differed from one another. However, following chronic treatment with LSD (post-tx), animals tested on LSD showed a significant reduction in their percent responding on the LSD lever relative to either their pretreatment choice behavior or relative to rats treated chronically with vehicle and tested on LSD.

Demonstration of development of behavioral tolerance to repeated LSD in a drug discrimination paradigm. Prior to chronic treatment (pre-tx; solid bars), neither the two groups of rats tested on vehicle nor the two groups tested on LSD (60 μg/kg) differed from one another. At 24 h following chronic treatment with LSD (130 μg/kg s.c. LSD for 5 days: post-tx; hatched bars), the choice behavior of animals tested on LSD (60 μg) was significantly different from pretreatment choice behavior.
Effect of Chronic LSD Treatment on LSD-Stimulated [35S]GTPγS Binding
To determine the effect of chronic LSD treatment on subsequent serotonin receptor function, receptor-stimulated [35S]GTPγS binding was examined. LSD-stimulated [35S]GTPγS binding was quantified in medial prefrontal cortex (mPFC), prefrontal cortex (PFC), frontal parietal cortex-motor (FPC-M), frontal parietal cortex-somatosensory (FPC-S), and Cl in brain sections from rats treated for 5 days with vehicle or LSD (130 μg/kg s.c.). Representative autoradiograms of the binding of [35S]GTPγS at the level of mPFC (+2.7 mm from bregma) are shown in Figure 3. Incubation of the section with 5.0 nM LSD significantly increased [35S]GTPγS binding in all of the brain regions from both vehicle- (Veh) and LSD-treated rats (Table 1). However, in the mPFC, the increase in [35S]GTPγS binding was significantly attenuated after chronic LSD (Table 1). LSD-stimulated [35S]GTPγS binding was also quantified in anterior cingulate cortex (ACC), FPC-M, FPC-S, endopiriform nucleus (En), STR, nucleus accumbens-core (nA-C), nucleus accumbens-shell (nA-S), and septum (S); representative autoradiograms of the binding of [35S]GTPγS at the level +1.2 mm from bregma are shown in Figure 4. LSD significantly increased [35S]GTPγS binding in ACC, FPC-M, FPC-S, En, and S, but no increase in [35S]GTPγS binding was found in STR, nA-C, and nA-S (Table 2). In the ACC, as in the mPFC, the increase in [35S]GTPγS binding was significantly attenuated after chronic LSD (Table 2).

Representative autoradiograms of [35S]GTPγS binding in sections of rat brain plate +2.7 mm from the bregma (Paxinos and Watson, 1986). Experimental conditions are identified by chronic treatment condition/drug incubation. (a) Veh/LSD, (b) Veh/Veh, (c) LSD/LSD, (d) LSD/Veh. Basal [35S]GTPγS binding appears to be similar between rats that were treated for 5 days with vehicle (panel b) or LSD (130 μg/kg s.c. for 5 days; panel d). LSD (5 nM) stimulated an increase in [35S]GTPγS binding in the mPFC, PFC, FPC-M, FPC-S, and Cl (panel a). After chronic LSD, the LSD-induced [35S]GTPγS binding appears to be reduced (panel c). For group data, see Table 1.

Representative autoradiograms of [35S]GTPγS binding in sections of rat brain plate +1.2 mm from the bregma (Paxinos and Watson, 1986). Experimental conditions are identified by chronic treatment condition/drug incubation: (a) Veh/LSD, (b) Veh/Veh, (c) LSD/LSD, (d) LSD/Veh. Basal [35S]GTPγS binding was similar between rats that were treated for 5 days with vehicle (panel b) or LSD (130 μg/kg s.c. for 5 days; panel d); LSD (5 nM) stimulated an apparent increase in [35S]GTPγS binding in the ACC, FPC-M, FPC-S, En, and S (panel a). The LSD-induced [35S]GTPγS binding appears to be reduced in the ACC from rats receiving repeated LSD (panel c). For group data, see Table 2.
Effect of Chronic LSD Treatment on DOI-Stimulated [35S]GTPγS Binding
In order to determine the relative contribution of 5-HT2A or 5-HT2C receptors to the reduced [35S]GTPγS binding in the mPFC and ACC, we used the selective 5-HT2A and 5-HT2C receptor agonist DOI along with the selective 5-HT2A receptor antagonist MDL 100907 and the selective 5-HT2C receptor antagonist SB206553. Representative autoradiograms of the binding of DOI-induced [35S]GTPγS binding are shown in Figures 5 and 6. Incubation of the section with 5.0 nM DOI significantly increased [35S]GTPγS binding in mPFC and ACC in rat sections obtained from both Veh- and LSD-treated rats (Tables 3 and 4). In both the mPFC (Table 3) and ACC (Table 4), the increase in DOI-stimulated [35S]GTPγS binding was significantly attenuated after chronic LSD. The DOI-stimulated increase in [35S]GTPγS binding in mPFC and ACC from both vehicle- and LSD-treated rats was blocked by the 5-HT2A receptor antagonist MDL 100907 (10 nM) but not the 5-HT2C receptor antagonist SB206553 (10 nM).

In order to test the contribution of 5-HT2A and/or 5-HT2C receptors in [35S]GTPγS binding, the 5-HT2A and 5-HT2C receptor agonist DOI was used as the stimulating compound. Representative autoradiograms of [35S]GTPγS binding in sections of rat brain plate +2.7 mm from bregma are shown (Paxinos and Watson, 1986). Basal [35S]GTPγS binding was similar between rats that were treated for 5 days with vehicle and those treated with LSD (130 μg/kg s.c. for 5 days). DOI (5 nM) stimulated an increase in [35S]GTPγS binding in mPFC; however, the DOI-induced [35S]GTPγS binding appears to be reduced after chronic LSD. The DOI-stimulated increase in [35S]GTPγS binding in mPFC from both vehicle- and LSD-treated rats was blocked by the 5-HT2A receptor antagonist MDL 100907 (10 nM) but not the 5-HT2C receptor antagonist SB206553 (10 nM). For group data, see Table 3.

Representative autoradiograms of [35S]GTPγS binding in sections of rat brain +1.2 mm from bregma (Paxinos and Watson, 1986). Basal [35S]GTPγS binding was similar between rats that were treated for 5 days with vehicle and those treated with LSD (130 μg/kg s.c. for 5 days). DOI (5 nM) stimulated an increase in [35S]GTPγS binding in ACC; however, the DOI-induced [35S]GTPγS binding appears to be reduced after chronic LSD. The DOI-stimulated increase in [35S]GTPγS binding in ACC from both vehicle- and LSD-treated rats was blocked by the 5-HT2A receptor antagonist MDL 100907 (10 nM) but not the 5-HT2C receptor antagonist SB206553 (10 nM). For group data, see Table 4.
Effect of Chronic LSD on 125I-LSD Binding
Chronic administration of 130 μg/kg LSD for 5 days resulted in a decrease in cortical 5-HT2A receptor binding. Representative autoradiograms of the binding for vehicle-and LSD-treated rats at the level +2.7 mm from bregma are shown in Figure 7. Treatment of rats with LSD for 5 days resulted in a significant decrease in 125I-LSD binding to 5-HT2A receptors in ACC, Cl, En, FPC-M, FPC-SS, mPFC, and PFC (Table 5).

Representative autoradiograms of 125I-LSD binding in sections of rat brain plate +2.7 mm from bregma (Paxinos and Watson, 1986). Rats were treated for 5 days with vehicle or LSD (130 μg/kg s.c. for 5 days). Sections were incubated with 125I-LSD (0.5 nM) in the presence of 1 μM sulpiride to mask dopamine D2 receptors and with 1 μM SB206553 to mask 5-HT2C receptors. Nonspecific binding was defined by the addition of 30 μM methysergide. For group data, see Table 5.
DISCUSSION
Extensive evidence convincingly demonstrates a key role for 5-HT2A receptors in hallucinogen-induced behavioral effects, including recent data in normal human subjects (Vollenweider et al, 1998). However, the precise brain site of action of LSD has not been elucidated. The current paper utilizes a functional neuroanatomical approach to evaluate the role of 5-HT2A receptors in specific brain sites. We asked the question, do anatomically localized changes in 5-HT2A receptors occur in concert with behavioral tolerance? Behavioral tolerance was evaluated in rats trained to discriminate 60 μg/kg LSD from saline in a two-lever drug discrimination paradigm. The drug discrimination paradigm is a powerful and sensitive method to evaluate subjective effects of drugs. This behavior in laboratory animals parallels the drug effects described by humans (Altman et al, 1976; Appel, 1990). To produce tolerance, rats were treated daily for 5 days with 130 μg/kg of LSD (as described by Buckholtz et al, 1990), and then tested for choice behavior in drug discrimination 24 h later. Rats treated chronically with LSD showed a significant 44% reduction in LSD lever responding (from 91 to 51%), while rats treated chronically with saline showed no change in LSD lever responding, indicating tolerance development. Previous studies have demonstrated tolerance to other behavioral effects of LSD in rats. LSD disruptive effects on rope climbing (Freedman et al, 1958) and fixed ratio lever pressing (Freedman et al, 1964; Winter, 1971) diminish after repeated administration, as does LSD-induced neophobia, a measure of avoidance of a novel environment (Adams and Geyer, 1985). Humans also become tolerant to the effects of LSD; subjects receiving multiple doses of LSD develop tolerance to the mood-altering effects, as well as the elementary and true hallucinations, perceptual distortions, and feelings of unreality and depersonalization (Isbell et al, 1956). The present experiments demonstrating a reduction in the stimulus intensity of LSD after chronic high doses of LSD are in agreement with a previous report that rats receiving chronic DOI show a marked decrease in percent DOI lever selection when tested with a challenge dose of DOI (Smith et al, 1999).
In parallel to behavioral tolerance in the LSD discrimination paradigm, we observed a significant reduction in LSD-induced [35S]GTPγS turnover after repeated LSD treatment. [35S]GTPγS autoradiography is a method that allows visualization and quantification of functional G-protein-coupled receptors in brain slices (Sim et al, 1997a). This method has been used, for example, to demonstrate changes in μ-opioid receptor function after chronic morphine and heroin treatments (Sim et al, 1996b; Sim-Selley et al, 2000), decreases in cannabinoid receptor activation after repeated tetrahydrocannabinol (Sim et al, 1996a), and heterologous regulation of 5-HT1A receptors (Valdez et al, 2002). In the current study, autoradiography of LSD-stimulated [35S]GTPγS binding revealed a region-specific attenuation of the cell signaling capacity of 5-HT2A receptors in rats chronically treated with LSD. Of the 11 areas examined, the decrease in LSD-stimulated [35S]GTPγS binding was localized to the mPFC and ACC. Since LSD interacts with many receptors, we performed a follow-up experiment to determine the contribution of 5-HT2A or 5-HT2C receptors using DOI, a more selective 5-HT2A/2C agonist. We confirmed the finding first reported by Adlersberg et al (2000) that DOI produces a significant increase in [35S]GTPγS binding in the frontal cortex. Furthermore, DOI-promoted [35S]GTPγS binding in mPFC and ACC was blocked by the 5-HT2A selective antagonist MDL100907, but not by the 5-HT2C receptor antagonist SB206553, demonstrating that the signal is mediated by 5-HT2A receptors. As was found for LSD, there was a significant reduction in DOI-induced [35S]GTPγS binding in the ACC and mPFC after chronic LSD treatment, indicating neuroadaptive changes in 5-HT2A receptors after LSD treatment that correspond with the expression of behavioral tolerance to LSD.
The findings from this study show that behavioral tolerance to the cue properties of LSD after chronic administration is associated with decreased 5-HT2A receptor signaling in the mPFC and ACC. The 5-HT2A receptor is widely expressed throughout the rat brain, including cerebral cortex, hippocampus, basal ganglia, amygdala, cerebellum, brain stem, and spinal cord (Mengod et al, 1990; Pompeiano et al, 1994; Cornea-Hebert et al, 1999). Areas with high expression include the mPFC and ACC where the receptor is located throughout the cortical layers with the densest expression on layer V pyramidal cells (Willins et al, 1997; Cornea-Hebert et al, 1999). Indeed, our agonist-stimulated [35S]GTPγS binding demonstrated a visible increase in G-protein activation in cortical layers IV/V (see Figures 3, 4, 5 and 6). In addition, our laboratory and others have demonstrated that acute LSD administration produces a robust increase in c-fos protein expression in the mPFC and ACC that is mediated by 5-HT2A receptors (Frankel and Cunningham, 2002; Gresch et al, 2002). Taken together, these findings provide converging behavioral and biochemical evidence that 5-HT2A receptors in ACC and mPFC are key sites in the action of LSD. Interestingly, in normal healthy human volunteers, the hallucinogen psilocybin elicits an increase in glucose metabolism in frontomedial and frontolateral cortex, ACC, and temporomedial cortex (Vollenweider et al, 1997), a pattern consistent with the present data.
In order to determine whether the reductions in [35S]GTPγS binding were due to changes downstream of the receptor in receptor–G-protein coupling efficiency or due to an alteration in receptor density, we performed 5-HT2A receptor autoradiography. The binding of 125I-LSD-labeled 5-HT2A receptors in cortical areas was decreased after chronic LSD treatment, consistent with a previous finding using dissected brain regions in a membrane binding assay (Buckholtz et al, 1990). In tolerant animals, 5-HT2A receptor binding sites and functional 5-HT2A receptors were both decreased in mPFC and ACC, but not in STR or nucleus accumbens. Thus, the reduction in 5-HT2A-mediated [35S]GTPγS binding after chronic LSD treatment appears to be explained by a reduction in 5-HT2A receptor number; however, a decrease in receptor number did not always translate into a reduced downstream signal. 125I-LSD-labeled 5-HT2A receptors were reduced in other cortical areas where there were no significant changes in the measure of functional receptor. These discrepancies may relate to the sensitivity of the assays; the [35S]GTPγS binding assay is hampered by a large basal signal, which may reflect endogenous constitutively active GPCRs. However, it is also possible that there are regional differences in the degree of receptor reserve that explain the functional differences. These results demonstrate the importance of measuring both the functional state of the receptors and the density of receptors after chronic drug treatment.
Interestingly, the magnitude and regional specificity of agonist-induced [35S]GTPγS binding differed between sections treated with LSD and DOI. In addition to the mPFC and ACC, LSD, but not DOI, produced a significant signal in motor and somatosensory parietal cortex, En, and septum. Studies by Adlersberg et al (2000) and our own unpublished work suggest that the concentration (5 nM) of LSD and DOI used in the assay is at or near maximum, suggesting that these two agonists may differ in their efficacy in the [35S]GTPγS binding assay, thus producing differing activation patterns. An alternative possibility for these differences relates to drug specificity; LSD interacts with multiple receptors, including most of the serotonin family as well as dopamine and adrenergic receptors, compared with DOI, which is selective for 5-HT2A and 5-HT2C receptors. Thus, the [35S]GTPγS signal produced by LSD may include other activated G-protein-coupled receptors in addition to the 5-HT2 receptors. For example, the septum, which contains a high concentration of 5-HT1A receptors (Sim et al, 1997b), had a robust LSD-induced [35S]GTPγS signal, but this region was unresponsive to DOI incubation. The possibility that the LSD signal in septum includes a 5-HT1A receptor component is consistent with the finding that the 5-HT1A receptor agonist 8-OH-DPAT increases septal [35S]GTPγS binding (Sim et al, 1997b). The LSD-induced [35S]GTPγS binding in the septum was unaltered by chronic LSD treatment, suggesting that 5-HT1A receptor signaling in the septum was not reduced after repeated LSD treatment. This finding is consistent with the lack of 5-HT1A receptor downregulation after chronic LSD (Buckholtz et al, 1990).
The findings from the current study further support the hypothesis that 5-HT2A receptors mediate the behavioral effects of hallucinogens. Previous studies have demonstrated that the discriminative stimulus properties of LSD are blocked by selective 5-HT2A receptor antagonists. To extend these findings, we have demonstrated a reduction in 5-HT2A receptor signaling in the mPFC and ACC, accompanying attenuation of the LSD discriminative cue in a chronic treatment paradigm that elicits tolerance. Furthermore, our demonstration of reduced LSD-mediated signaling in the mPFC and ACC indicates that a significant component of the discriminative stimulus is mediated within these brain sites. Aghajanian and Marek proposed that hallucinogenic drugs interact with 5-HT2A receptors on thalamocortical terminals in the mPFC and parietal cortex to augment glutamate neurotransmission (Marek and Aghajanian, 1998; Aghajanian and Marek, 2000). The current results suggest that chronic LSD exposure could modulate this regulatory process, resulting in altered activity of cortical pyramidal cells. Furthermore, a functional consequence of decreased 5-HT2A receptors after chronic hallucinogens may be impairment in cognitive processing (Williams et al, 2002; Passetti et al, 2003).
References
Adams LM, Geyer MA (1985). A proposed animal model for hallucinogens based on LSD's effects on patterns of exploration in rats. Behav Neurosci 99: 881–900.
Adlersberg M, Arango V, Hsiung S, Mann JJ, Underwood MD, Liu K et al (2000). In vitro autoradiography of serotonin 5-HT(2A/2C) receptor-activated G protein: guanosine-5′-(gamma-[(35)S]thio)triphosphate binding in rat brain. J Neurosci Res 61: 674–685.
Aghajanian GK, Marek GJ (2000). Serotonin model of schizophrenia: emerging role of glutamate mechanisms. Brain Res Brain Res Rev 31: 302–312.
Altman JL, Albert JM, Milstein SL, Greenberg I (1976). Drugs as the discriminative events in humans. Psychopharmacol Commun 2: 327–330.
Appel JB (1990). Drug discrimination as a tool in drug abuse research. Psychopharmacology 101: S65.
Appel JB, West WB, Buggy J (2004). LSD, 5-HT (serotonin), and the evolution of a behavioral assay. Neurosci Biobehav Rev 27: 693–701.
Buckholtz NS, Zhou DF, Freedman DX, Potter WZ (1990). Lysergic acid diethylamide (LSD) administration selectively downregulates serotonin2 receptors in rat brain. Neuropsychopharmacology 3: 137–148.
Callahan PM, Appel JB (1988). Differences in the stimulus properties of 3, 4-methylenedioxyamphetamine and 3, 4- methylenedioxymethamphetamine in animals trained to discriminate hallucinogens from saline. J Pharmacol Exp Ther 246: 866–870.
Colpaert FC, Meert TF, Niemegeers CJ, Janssen PA (1985). Behavioral and 5-HT antagonist effects of ritanserin: a pure and selective antagonist of LSD discrimination in rat. Psychopharmacology 86: 45–54.
Cornea-Hebert V, Riad M, Wu C, Singh SK, Descarries L (1999). Cellular and subcellular distribution of the serotonin 5-HT2A receptor in the central nervous system of adult rat. J Comp Neurol 409: 187–209.
Cunningham KA, Appel JB (1987). Neuropharmacological reassessment of the discriminative stimulus properties of d-lysergic acid diethylamide (LSD). Psychopharmacology 91: 67–73.
Fiorella D, Rabin RA, Winter JC (1995). Role of 5-HT2A and 5-HT2C receptors in the stimulus effects of hallucinogenic drugs. II: reassessment of LSD false positives. Psychopharmacology (Berl) 121: 357–363.
Frankel PS, Cunningham KA (2002). The hallucinogen d-lysergic acid diethylamide (d-LSD) induces the immediate-early gene c-Fos in rat forebrain. Brain Res 958: 251–260.
Freedman DX, Aghajanian GK, Ornitz EM (1958). Patterns of tolerance to lysergic acid diethylamide and mescaline in rats. Science 127: 1173–1174.
Freedman DX, Appel JB, Hartman FR, Molliver ME (1964). Tolerance to behavioral effects of LSD-25 in rat. J Pharmacol Exp Ther 143: 309–313.
Glennon RA, Hauck AE (1985). Mechanistic studies on DOM as a discriminative stimulus. Pharmacol Biochem Behav 23: 937–941.
Glennon RA, Titeler M, McKenney JD (1984). Evidence for 5-HT2 involvement in the mechanism of action of hallucinogenic agents. Life Sci 35: 2505–2511.
Gresch PJ, Strickland LV, Sanders-Bush E (2002). Lysergic acid diethylamide-induced Fos expression in rat brain: role of serotonin-2A receptors. Neuroscience 114: 707–713.
Isbell H, Belleville RE, Fraser HF, Wikler A, Logan CR (1956). Studies on lysergic acid diethylamide (LSD-25). I. Effects in former morphine addicts and development of tolerance during chronic intoxication. AMA Arch Neurol Psychiatry 76: 468–478.
Laitinen JT (1999). Selective detection of adenosine A1 receptor-dependent G-protein activity in basal and stimulated conditions of rat brain [35S]guanosine 5′-(gamma-thio)triphosphate autoradiography. Neuroscience 90: 1265–1279.
Leysen JE, Janssen PF, Niemegeers CJ (1989). Rapid desensitization and down-regulation of 5-HT2 receptors by DOM treatment. Eur J Pharmacol 163: 145–149.
Marek GJ, Aghajanian GK (1998). Indoleamine and the phenethylamine hallucinogens: mechanisms of psychotomimetic action. Drug Alcohol Depend 51: 189–198.
Mengod G, Pompeiano M, Martinez Mir MI, Palacios JM (1990). Localization of the mRNA for the 5-HT2 receptor by in situ hybridization histochemistry. Correlation with the distribution of receptor sites. Brain Res 524: 139–143.
Passetti F, Dalley JW, Robbins TW (2003). Double dissociation of serotonergic and dopaminergic mechanisms on attentional performance using a rodent five-choice reaction time task. Psychopharmacology (Berl) 165: 136–145.
Paxinos G, Watson C (1986). The Rat Brain in Stereotaxic Coordinates. Academic Press: New York.
Pompeiano M, Palacios JM, Mengod G (1994). Distribution of the serotonin 5-HT2 receptor family mRNAs: comparison between 5-HT2A and 5-HT2C. Mol Brain Res 23: 163–178.
Sim LJ, Hampson RE, Deadwyler SA, Childers SR (1996a). Effects of chronic treatment with delta9-tetrahydrocannabinol on cannabinoid-stimulated [35S]GTPgammaS autoradiography in rat brain. J Neurosci 16: 8057–8066.
Sim LJ, Selley DE, Childers SR (1997a). Autoradiographic visualization in brain of receptor-G protein coupling using [35S]GTP gamma S binding. Methods Mol Biol 83: 117–132.
Sim LJ, Selley DE, Dworkin SI, Childers SR (1996b). Effects of chronic morphine administration on mu opioid receptor-stimulated [35S]GTPgammaS autoradiography in rat brain. J Neurosci 16: 2684–2692.
Sim LJ, Xiao R, Childers SR (1997b). In vitro autoradiographic localization of 5-HT1A receptor-activated G-proteins in the rat brain. Brain Res Bull 44: 39–45.
Sim-Selley LJ, Selley DE, Vogt LJ, Childers SR, Martin TJ (2000). Chronic heroin self-administration desensitizes mu opioid receptor-activated G-proteins in specific regions of rat brain. J Neurosci 20: 4555–4562.
Smith RL, Barrett RJ, Sanders-Bush E (1995). Neurochemical and behavioral evidence that quipazine–ketanserin discrimination is mediated by serotonin2A receptor. J Pharmacol Exp Ther 275: 1050–1057.
Smith RL, Barrett RJ, Sanders-Bush E (1999). Mechanism of tolerance development to 2, 5-dimethoxy-4-iodoamphetamine in rats: down-regulation of the 5-HT2A, but not 5-HT2C, receptor. Psychopharmacology (Berl) 144: 248–254.
Titeler M, Lyon RA, Glennon RA (1988). Radioligand binding evidence implicates the brain 5-HT2 receptor as a site of action for LSD and phenylisopropylamine hallucinogens. Psychopharmacology 94: 213–216.
Valdez M, Burke TF, Hensler JG (2002). Selective heterologous regulation of 5-HT1A receptor-stimulated 35S GTPgammaS binding in the anterior cingulate cortex as a result of 5-HT2 receptor activation. Brain Res 957: 174–182.
Vollenweider FX, Leenders KL, Scharfetter C, Maguire P, Stadelmann O, Angst J (1997). Positron emission tomography and fluorodeoxyglucose studies of metabolic hyperfrontality and psychopathology in the psilocybin model of psychosis. Neuropsychopharmacology 16: 357–372.
Vollenweider FX, Vollenweider-Scherpenhuyzen MF, Babler A, Vogel H, Hell D (1998). Psilocybin induces schizophrenia-like psychosis in humans via a serotonin-2 agonist action. Neuroreport 9: 3897–3902.
Williams GV, Rao SG, Goldman-Rakic PS (2002). The physiological role of 5-HT2A receptors in working memory. J Neurosci 22: 2843–2854.
Willins DL, Deutch AY, Roth BL (1997). Serotonin 5-HT2A receptors are expressed on pyramidal cells and interneurons in the rat cortex. Synapse 27: 79–82.
Winter JC (1971). Tolerance to a behavioral effect of lysergic acid diethylamide and cross-tolerance to mescaline in the rat: absence of a metabolic component. J Pharmacol Exp Ther 178: 625–630.
Acknowledgements
This work was supported by research grants from National Institute of Drug Abuse DA05181 (ESB), DA15165 (PJG), and Veterans Administration Medical Center (RJR). We thank Kathleen Patterson for her expert technical assistance. Preliminary results from these studies were presented at the 33rd Annual Meeting of the Society for Neuroscience, New Orleans, LA, 2003.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gresch, P., Smith, R., Barrett, R. et al. Behavioral Tolerance to Lysergic Acid Diethylamide is Associated with Reduced Serotonin-2A Receptor Signaling in Rat Cortex. Neuropsychopharmacol 30, 1693–1702 (2005). https://doi.org/10.1038/sj.npp.1300711
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1300711
Keywords
This article is cited by
-
Assessing the risk–benefit profile of classical psychedelics: a clinical review of second-wave psychedelic research
Psychopharmacology (2022)
-
Pharmacology of ayahuasca administered in two repeated doses
Psychopharmacology (2012)
-
The neurobiology of psychedelic drugs: implications for the treatment of mood disorders
Nature Reviews Neuroscience (2010)
-
Hallucinogene paddenstoelen
Verslaving (2010)
-
Enduring Deficits in Sustained Visual Attention during Withdrawal of Intravenous Methylenedioxymethamphetamine Self-Administration in Rats: Results from a Comparative Study with d-Amphetamine and Methamphetamine
Neuropsychopharmacology (2007)
