
Abstract
During heart transplantation, storage in cold preservation solution is thought to protect the organ by slowing metabolism, providing osmotic support and minimizing ischaemia–reperfusion (IR) injury following transplantation in the recipient1,2. Despite its widespread use, our understanding of the metabolic changes prevented by cold storage and how warm ischaemia leads to damage is surprisingly poor. Here, we compare the metabolic changes during warm ischaemia (WI) and cold ischaemia (CI) in mouse, pig and human hearts. We identify common metabolic alterations during WI and CI, thereby elucidating mechanisms underlying the benefits of CI and how WI causes damage. Succinate accumulation is a major feature within ischaemic hearts across species, and CI slows succinate generation, thereby reducing tissue damage upon reperfusion caused by the production of mitochondrial reactive oxygen species (ROS)3,4. Importantly, the inevitable periods of WI during organ procurement lead to the accumulation of damaging levels of succinate during transplantation, despite cooling organs as rapidly as possible. This damage is ameliorated by metabolic inhibitors that prevent succinate accumulation and oxidation. Our findings suggest how WI and CI contribute to transplant outcome and indicate new therapies for improving the quality of transplanted organs.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
Data availability
Metabolomic data have been uploaded as study MTBLS1085 to MetaboLights (https://www.ebi.ac.uk/metabolights). The original synthetic chemistry data have been deposited at https://doi.org/10.5525/gla.researchdata.646. Other data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Jahania, M. S., Sanchez, J. A., Narayan, P., Lasley, R. D. & Mentzer, R. M. Jr. Heart preservation for transplantation: principles and strategies. Ann. Thorac. Surg. 68, 1983–1987 (1999).
Southard, J. H. & Belzer, F. O. Organ preservation. Annu. Rev. Med. 46, 235–247 (1995).
Chouchani, E. T. et al. A unifying mechanism for mitochondrial superoxide production during ischemia-reperfusion injury. Cell Metab. 23, 254–263 (2016).
Chouchani, E. T. et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 515, 431–435 (2014).
Pagani, F. D. Use of heart donors following circulatory death: a viable addition to the heart donor pool. J. Am. Coll. Cardiol. 73, 1460–1462 (2019).
Johnson, R. J., Bradbury, L. L., Martin, K. & Neuberger, J. Organ donation and transplantation in the UK-the last decade: a report from the UK national transplant registry. Transplantation 97, S1–S27 (2014).
Chew, H. C. et al. Outcomes of donation after circulatory death heart transplantation in australia. J. Am. Coll. Cardiol. 73, 1447–1459 (2019).
Dhital, K. K. et al. Adult heart transplantation with distant procurement and ex-vivo preservation of donor hearts after circulatory death: a case series. Lancet 385, 2585–2591 (2015).
Nasralla, D. et al. A randomized trial of normothermic preservation in liver transplantation. Nature 557, 50–56 (2018).
Coffey, J. C. et al. The influence of functional warm ischemia time on DCD liver transplant recipients’ outcomes. Clin. Transplant. 31, e13068 (2017).
Blok, J. J. et al. Longterm results of liver transplantation from donation after circulatory death. Liver Transplant. 22, 1107–1114 (2016).
Eltzschig, H. K. & Eckle, T. Ischemia and reperfusion—from mechanism to translation. Nat. Med. 17, 1391–1401 (2011).
Liu, F. & Kang, S. M. Heterotopic heart transplantation in mice. J. Vis. Exp. 6, 238 (2007).
Niimi, M. The technique for heterotopic cardiac transplantation in mice: experience of 3000 operations by one surgeon. J. Heart Lung Transplant. 20, 1123–1128 (2001).
Zhang, Q. et al. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature 464, 104–107 (2010).
Nakahira, K. et al. Circulating mitochondrial DNA in patients in the ICU as a marker of mortality: derivation and validation. PLoS Med. 10, e1001577 (2013).
Ehinger, J. K. et al. Cell-permeable succinate prodrugs bypass mitochondrial complex I deficiency. Nat. Commun. 7, 12317 (2016).
Valls-Lacalle, L. et al. Selective inhibition of succinate dehydrogenase in reperfused myocardium with intracoronary malonate reduces infarct size. Sci. Rep. 8, 2442 (2018).
Valls-Lacalle, L. et al. Succinate dehydrogenase inhibition with malonate during reperfusion reduces infarct size by preventing mitochondrial permeability transition. Cardiovasc. Res. 109, 374–384 (2016).
Bundgaard, A. et al. Metabolic adaptations during extreme anoxia in the turtle heart and their implications for ischemia-reperfusion injury. Sci. Rep. 9, 2850 (2019).
Zhang, J. et al. Accumulation of succinate in cardiac ischemia primarily occurs via canonical krebs cycle activity. Cell Rep. 23, 2617–2628 (2018).
Dare, A. J. et al. The mitochondria-targeted anti-oxidant MitoQ decreases ischemia-reperfusion injury in a murine syngeneic heart transplant model. J. Heart Lung Transplant. 34, 1471–1480 (2015).
Mackay, G. M., Zheng, L., van den Broek, N. J. & Gottlieb, E. Analysis of cell metabolism using LC-MS and isotope tracers. Meth. Enzymol. 561, 171–196 (2015).
Strehler, B. L. in Methods in Enzymatic Analysis (ed. Bergmeyer, U.) 2112–2126 (Academic Press, 1974).
Passonneau, J. V. & Lauderdale, V. R. A comparison of three methods of glycogen measurement in tissues. Anal. Biochem. 60, 405–412 (1974).
Santos, J. H., Meyer, J. N., Mandavilli, B. S. & Van Houten, B. Quantitative PCR-based measurement of nuclear and mitochondrial DNA damage and repair in mammalian cells. Methods Mol. Biol. 314, 183–199 (2006).
Ritchie, M. E. et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nuc. Acids Res. 43, e47 (2015).
Acknowledgements
Work in the M.P.M. laboratory was supported by the Medical Research Council UK (MC_U105663142) and by a Wellcome Trust Investigator award (110159/Z/15/Z) to M.P.M. Work in the C.F. laboratory was supported by the Medical Research Council (MRC_MC_UU_12022/6). Work in the K.S.P. laboratory was supported by the Medical Research Council UK. Work in the RCH lab laboratory was supported by a Wellcome Trust Investigator award (110158/Z/15/Z) and a PhD studentship for .L.P from the University of Glasgow. A.V.G. was supported by a PhD studentship funded by the National Institute for Health Research Blood and Transplant Research Unit (NIHR BTRU) in Organ Donation and Transplantation at the University of Cambridge in collaboration with Newcastle University and in partnership with NHS Blood and Transplant (NHSBT). The views expressed are those of the author(s) and not necessarily those of the NHS, the NIHR, the Department of Health or NHSBT. We are grateful to the donors, the donor families, National Health Service Blood transfusion and Transplantation, and the Cambridge Biorepository for Translational Medicine for access to human samples.
Author information
Authors and Affiliations
Contributions
J.L.M., A.S.H.C., K.S.-P., R.C.H., T.K., M.H., K.M., C.F. and M.P.M. designed the experimental protocols. A.V.G., F.M.A., H.A.P., S.T.C., L.T., M.M.H., E.C.H., A.S.H.C., T.E.B., L.P., A.L., E.N. and J.L.M. performed the studies. J.L.M., A.S.H.C., A.M.J. and A.R. analysed the data. J.L.M., A.S.H.C., K.S.-P., C.F. and M.P.M. wrote the manuscript with assistance from all the other authors.
Corresponding author
Ethics declarations
Competing interests
M.P.M., T.K. and R.C.H. have submitted a patent application on the use of dimethyl malonate to prevent IR injury.
Additional information
Peer review information Primary Handling Editor: Ana Mateus.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
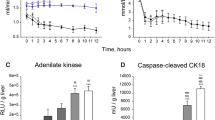
Extended Data Fig. 1 Effect of cardioplegia on ATP/ADP during WI in the mouse heart and cardiac AMP fold change during WI and CI.
a, ATP/ADP ratios of mouse hearts were assessed as described in Fig. 1 during WI ± cardioplegia at the onset of WI. Individual normoxic controls were used for WI ± cardioplegia, 6 min, n = 3; all others n = 4. Data are means ± SEM, WI, normoxic control and 12 min, and WI + cardioplegia, n = 3–4. b, AMP levels were determined in heart tissue after various times of WI or CI and are given as fold change relative to normoxic controls. Data are means ± s.e.m. Mouse normoxic controls, n = 11; CI, 240 min; n = 4; all others n = 5. n = 5 (pig), 4 (human).
Extended Data Fig. 2 Glycogen and glycolysis during WI and CI.
a, Glycogenolysis and glycolysis in ischaemia. G-6-P, glucose-6-phosphate; Gly-3P, glyceraldehyde-3 phosphate; BPG, bisphosphoglycerate; 3PG, 3-phosphoglycerate; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; LDH, lactate dehydrogenase; PEP, phosphoenolpyruvate. b,c, Heart glycogen during WI and CI in (b) mouse and (c) pig. Mean ± s.e.m. Pig, n = 3. Mouse, WI, 0, 6, 12, 30 min, n = 5; 120 min, n = 4; CI, 6, 12, 30, 60 min, n = 7; 120 and 240 min, n = 3. d–g, Heart lactate levels during WI and CI in (d,g) mouse, (e) pig and (f) human. Dashed red line shows when both glycogenolysis and lactate accumulation stop during WI. Data are shown as the fold-change relative to normoxic controls, mean ± SEM, n = 4. h, Heart lactate/pyruvate ion current ratio during WI and CI in the mouse, determined as fold change relative to normoxic controls. Data are relative changes in the ratio of the ion current for these metabolites. Mean ± s.e.m. Normoxic controls n = 11, WI, n = 5; CI, n = 4. i, Heart NADH/NAD++ ion current ratio during WI and CI in the mouse, shown as fold change relative to normoxic controls. Data are presented as relative changes in the ratio of the ion current for these metabolites, which is proportional to, but not the same as, the true ratio of the metabolite levels. Data mean ± s.e.m. Normoxic controls n = 11, WI, n = 5; CI, n = 4. NADH was not detectable in two control hearts and the lowest detectable value was then used.
Extended Data Fig. 3 Changes in glycolytic metabolites in the mouse heart during WI and CI.
Plots of the glycolytic metabolites detected in the mouse heart during WI and CI. Heart tissue was stored and analysed as described in Fig. 4. In most cases data are shown relative to normoxic control tissues. In some cases where the metabolite was undetectable in normoxic tissue the ion currents in arbitrary units (AU) are reported. Data are means ± s.e.m. For glucose -6-phosphate and fructose-6-phosphate, normoxic control, n = 11; 240 min, n = 4; rest, n = 5. For glyceraldehyde-3-phosphate, normoxic control, n = 11; 240 min WI, n = 4; 30 and 240 min CI, n = 3; rest, n = 5. Pyruvate was poorly detected, normoxic control, n = 11; WI, 6 min I, n = 4; 12 min, n = 1; 30 min, n = 2; 240 min, n = 4; CI, WI, 6 min, n = 5; 12 min, n = 4; 30 min, n = 2, 240 n = 1. For 3-phosphoglycerate and phosphoenol pyruvate ion counts were used, and for the normoxic control, n = 8. The ion counts for WI samples were either undetectable (no data point symbol) or n = 1 (data point sysmbol). For CI n = 3 for both compounds at 6 and 12 min, and n = 1 for 240 min. Phosphoenol pyruvate was not detected at 30 min and for 3-phosphoglycerate n = 1 at 30 min. F-1,6-BP, fructose 1,6-bisphosphate; 2-PG, 2-phosphoglycerate; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; AU, arbitrary units.
Extended Data Fig. 4 Metabolic changes during WI and CI.
Heat maps showing how 54 metabolites change in relative abundance during WI and CI in mouse, pig and human heart tissue compared to normoxic controls (Ct). Independent biological replicates for each condition are: mouse, Ct and WI 6 min, n = 6; CI, 480 min, n = 3; rest, n = 5. Pig, n = 5. Human, Ct, n = 5; rest, n = 4.
Extended Data Fig. 5 Extended set of metabolic changes during WI and CI in mice.
Heat map showing how 107 metabolites change in relative abundance in mouse hearts during WI and CI compared to normoxic controls (Ct). Independent biological replicates for each condition are: Ct, n = 6 and WI 6 min; 480 min CI, n = 3; rest, n = 5.
Extended Data Fig. 6 Time course of cardioplegia, flushing, temperature and succinate changes in the heart.
Succinate levels in mouse hearts were assessed as described in Fig. 3 during CI and WI ± cardioplegia at the onset of WI. Individual normoxic controls were used for CI and for WI ± cardioplegia. Data are means ± s.e.m. WI + cardioplegia, n = 3, rest n = 4. b, Mouse hearts were exposed and diastolic arrest was induced with St Thomas’ cardioplegia solution. Hearts were then rapidly excised and placed in storage medium at various temperatures, and after 12 min were rapidly . The succinate level was determined and is expressed relative to the amount in normoxic control hearts (data = means ± s.e.m., n = 4). There was a linear relationship between temperature and succinate accumulation (R2 = 0.998). c, Mouse hearts were rapidly excised as in b and placed in storage medium at 4 °C. The temperature of the apexes of the hearts were measured during cooling using a thermocouple placed into the apex of the heart through the ascending aorta. Mean ± s.e.m. (n = 3). d, Mouse hearts were retrieved by surgical preparation of the great vessels required for heterotopic heart transplantation and the temperature of the apex measured using a thermocouple. Diastolic arrest of mouse hearts was induced with cold St Thomas’ cardioplegia solution and topical cold saline was applied at the start of the retrieval process and then at regular intervals thereafter (blue arrows). The panel shows a typical result of three replicates. e, Succinate measured in hearts under WI, CI or standard retrieval (as described in d). Data are mean ± s.e.m., n = 5. f, Diacetoxymethyl succinate (AMS) administration and tissue succinate content. Mice were anaesthetized under isoflurane anaesthesia and a laparotomy was performed to expose the great vessels. The inferior vena cava (IVC) was infused with a bolus of 500 μl St Thomas’ cardioplegia solution and 5% DMSO ± 3.2 mg AMS. Immediately after administering the cardioplegia, the thorax of the mouse was opened and the heart rapidly removed and placed in ice cold PBS (this step took between 10–20 s). The hearts were stored in ice cold PBS for 30 min and then either clamp frozen in liquid nitrogen or were then incubated for a further 20 min at 37 °C in the carcass of the animal to mimic organ anastomosis and then were clamp frozen in liquid nitrogen. Absolute tissue succinate concentration was quantified by interpolation of the standard curve obtained from serial dilutions of commercially available standards that were run with the same batch of samples. Data mean of 4 hearts per conditions ± s.e.m. g, Effect of diacetoxymethyl fumarate (AMF) administration on cell levels of fumarate. C2C12 cells were incubated with 0, 0.25, 0.5 or 1 mM fumarate-AM diester for 30 min at 37 °C before extraction and analysis of fumarate by LC–MS/MS (mean ± s.e.m., n = 3 biological replicates). h,i, Comparison of AMF administration with that of AMS. Mice were anaesthetized under isoflurane anaesthesia and a laparotomy was performed to expose the great vessels. The inferior vena cava (IVC) was infused with a bolus of 500 μl St Thomas’ cardioplegia solution and 5% DMSO + 3.2 mg AMS or 30% DMSO + 3.18 mg AMF, which is equimolar with the dose of AMS used. Immediately after administering the cardioplegia, the thorax of the mouse was opened and the heart was rapidly removed and placed in ice-cold PBS (this step took between 10–20s). The hearts were stored in ice cold PBS for 30 min and then transplanted into a recipient as described in the legend to Fig. 4d. After 24 h, the plasma levels of troponin (h) and mtDNA relative to nuclear DNA (i) were assessed. Data are the mean of 4 hearts per conditions ± s.e.m. j,k, Hearts were infused with vehicle or DMM before being retrieved from the thorax as previously described. Hearts were then exposed to 12 min WI, followed by 30 min CI, then 20 min WI to mimic the changes during anastomosis. Hearts were clamp frozen before measuring malonate (j) and succinate (k) levels by LC–MS/MS (data are mean ± s.e.m., n = 3; *P < 0.05, **P < 0.001 by a two-tailed Student’s unpaired t-test).
Supplementary information
Supplementary Information
Supplementary Fig. 1 and Table 1
Supplementary Data 1
Tables of metabolites in mouse, pig and human hearts
Supplementary Data 2
Tables of metabolites in mouse, pig and human hearts
Rights and permissions
About this article

Cite this article
Martin, J.L., Costa, A.S.H., Gruszczyk, A.V. et al. Succinate accumulation drives ischaemia-reperfusion injury during organ transplantation. Nat Metab 1, 966–974 (2019). https://doi.org/10.1038/s42255-019-0115-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-019-0115-y
This article is cited by
-
Comprehensive bile acid pool analysis during ex-vivo liver perfusion in a porcine model of ischemia-reperfusion injury
Scientific Reports (2024)
-
Metabolite signaling in the heart
Nature Cardiovascular Research (2023)
-
Preventing mitochondrial reverse electron transport as a strategy for cardioprotection
Basic Research in Cardiology (2023)
-
Assessment of left ventricular tissue mitochondrial bioenergetics in patients with stable coronary artery disease
Nature Cardiovascular Research (2023)
-
Modeling energy depletion in rat livers using Nash equilibrium metabolic pathway analysis
Scientific Reports (2022)
