
Abstract
The history of the British Isles and Ireland is characterized by multiple periods of major cultural change, including the influential transformation after the end of Roman rule, which precipitated shifts in language, settlement patterns and material culture1. The extent to which migration from continental Europe mediated these transitions is a matter of long-standing debate2,3,4. Here we study genome-wide ancient DNA from 460 medieval northwestern Europeans—including 278 individuals from England—alongside archaeological data, to infer contemporary population dynamics. We identify a substantial increase of continental northern European ancestry in early medieval England, which is closely related to the early medieval and present-day inhabitants of Germany and Denmark, implying large-scale substantial migration across the North Sea into Britain during the Early Middle Ages. As a result, the individuals who we analysed from eastern England derived up to 76% of their ancestry from the continental North Sea zone, albeit with substantial regional variation and heterogeneity within sites. We show that women with immigrant ancestry were more often furnished with grave goods than women with local ancestry, whereas men with weapons were as likely not to be of immigrant ancestry. A comparison with present-day Britain indicates that subsequent demographic events reduced the fraction of continental northern European ancestry while introducing further ancestry components into the English gene pool, including substantial southwestern European ancestry most closely related to that seen in Iron Age France5,6.
Similar content being viewed by others


Main
The first millennium ce saw major demographic, cultural and political change in Europe, including the rise and fall of the Roman Empire, migration and the emergence of medieval institutions that shaped the modern world. The post-Roman transformation of lowland Britain was particularly profound. The end of the Roman administration in fifth century Britain preceded a dramatic shift in material culture, architecture, manufacturing and agricultural practice, and was accompanied by language change1. The archaeological record and place names indicate shared cultural features across the North Sea zone, in particular, along the east and southeast coasts of present-day England, Schleswig-Holstein and Lower Saxony (Germany), Frisia (Netherlands) and the Jutland peninsula (Denmark)2,3,4. Examples include the appearance of Grubenhäuser (sunken feature buildings), large cremation cemeteries and the styles of cremation urns or objects that used animal art and chip-carved metal7,8,9,10,11. Moreover, wrist clasps, as well as cruciform and square-headed brooches, found in sixth and seventh century Britain had attested southern Scandinavian origins12,13. Despite these similarities across the North Sea zone, there was also insular material culture that had no continental equivalent14,15. Adding to this, some places and geographical features such as rivers retained names of Celtic or late Latin origin16,17.
From the Renaissance to the present day, the primary explanatory narrative for these changes has been invasion and conquest followed by resettlement from the continent18. On the basis of a small set of written sources, it was supposed that the local Romano-British population was largely replaced by migrants from the Germanic-speaking part of the continent. However, the extent to which these traditional cultural historical interpretations explain patterns of material culture or agree with the historical accounts has been questioned18. For example, historical sources going back to Bede (writing in the eight century) indicated Jutes as settlers in Kent. But, in an issue that became known as ‘the problem of the Jutes’19,20,21, this historically attested migration is difficult to determine from or reconcile with the archaeological record. Indeed, material culture elements found in Kent resemble those of contemporary Merovingian France and Alemannic (southern) Germany, rather than the rest of England or Denmark. Such discrepancies between the archaeological record and historical narratives could be argued to support a rejection of migration or invasion hypotheses, and this was the preferred theoretical position of many archaeologists from the 1960s onwards1,18,22. By that time, many scholars favoured a model of elite dominance involving small, mobile warbands and the acculturation of the local British population. However, the available isotopic and DNA evidence, even if hitherto small scale, suggests that immigrants were less wealthy and buried alongside locals23,24,25,26,27,28, which does not fit a model of elite influence that could explain the adoption of a West Germanic language with apparently minimal influence from Celtic or Latin29,30,31,32.
There is a history of addressing these questions using genetic data. After early attempts to use ancient genetic data failed33, researchers turned to studies based on present-day populations and uniparentally inherited markers, but still without reaching consensus. Work based on present-day Y chromosomes inferred 50–100% replacement of male lineages during the Early Middle Ages in eastern England34,35. More recently, the first genome-wide study of present-day British people concluded that immigrant continental northern European ancestry makes up less than 50% of the present-day southeastern English gene pool36. However, populations change over time through drift and gene flow, so present-day populations may be poor proxies for ancient groups of unknown genetic makeup. The feasibility of ancient DNA analyses to inform on population history in Britain was first demonstrated with the report of genome-wide ancient DNA (aDNA) data26,37 from 20 individuals from the Iron Age to the Early Middle Ages, two studies that have provided unambiguous evidence for continental ancestry in early and middle Anglo-Saxon contexts.
Here we investigate early medieval population dynamics in England and across the North Sea zone with the first large-scale genome-wide study of aDNA in this time period and region, increasing the archaeogenetic record in England specifically, from 8 to 285 individuals. We target a comprehensive time transect of sites in the south and east of England, spanning predominantly the time period 450–850 ce, starting with early Anglo-Saxon cemeteries including Apple Down, Dover Buckland, Eastry, Ely, Hatherdene Close, Lakenheath, Oakington, Polhill and West Heslerton. This allows us to address questions concerning the extent of continental migration to England, and its effect on the local insular gene pool. In addition, the association of artefacts with individuals allows us to study the dynamics of the migration process at the community level.
New aDNA data
We sampled skeletal remains from 494 ancient northwestern Europeans from 37 different sites in England, Ireland, the Netherlands, Germany and Denmark, dated between approximately 200 and 1300 ce (Supplementary Note 1 and Supplementary Table 1). We prepared powder from skeletal material, extracted aDNA and converted it into double-stranded or single-stranded libraries (Methods). We selected 439 libraries for hybridization DNA capture to enrich for sequences that overlapped 1.24 million single-nucleotide polymorphisms (SNPs). For 40 libraries, we generated complete genomes without capture, with a mean coverage of 0.9×.
After quality filtering (Methods) and exclusion of duplicate individuals, genome-wide data for 460 individuals were available for analysis. These include 278 ancient individuals from England, and 182 individuals from neighbouring ancient populations in Ireland and the European continent (Fig. 1). We combined our newly reported data with published aDNA from 4,336 individuals (Supplementary Note 2), including 1,098 post-Neolithic genomes from northwestern Europe26,37,38,39,40,41,42,43,44. We also compiled a reference dataset of 10,176 present-day European individuals36,45,46,47 genotyped on an intersection of 445,171 SNPs (Supplementary Note 2). To aid interpretation of our genetic data, we also radiocarbon-dated 57 samples selected on the basis of ancestry composition, burial assemblage and preservation.

a, Spatial distribution of sites analysed in this study. b, Temporal distribution of samples analysed in this study, with site occupancy ranges indicated by bars. Non-transparent symbols indicate radiocarbon-dated samples; transparent symbols are scattered uniformly along site occupancy ranges. DK, Denmark; ENG, England; GER, Germany, IE, Ireland; NED, Netherlands.
Population shifts in post-Roman England
We performed principal component analysis (PCA) on 5,365 present-day northwestern Europeans, from Ireland to Sweden, and projected our ancient genomes onto this genetic variation (Fig. 2). For present-day variation, PC1 and PC2 broadly reflect geography, forming a V-shaped pattern from Scandinavians via individuals from northern Germany and the Netherlands towards those from Britain and Ireland. We highlight the position of individuals from present-day England (Fig. 2a), which follow a clinal distribution defined by the western British and Irish (WBI; which includes Irish, Northern Irish, Scottish and Welsh) at one extreme and overlapping present-day Dutch at the other extreme. The ancient genomes fall onto a slightly separate cline, with most of the early medieval individuals from Dutch, German and Danish sites plotting on top of present-day continental northern Europeans (CNEs; northern Germans and Danish), whereas Bronze and Iron Age individuals from Britain and Ireland cluster together with WBI (Fig. 2b). Of note, in contrast to the preceding Bronze and Iron Age individuals from Britain and Ireland, the majority of the early medieval samples from England (England EMA) plot together with the ancient individuals from the continental North Sea area along with the present-day CNEs. The divergence between prehistoric and early medieval individuals from England is also seen in the distribution of genetic distances (FST) as well as shared alleles (F4) on both the population (Extended Data Fig. 1) and the individual scale (Supplementary Fig. 3.3). We notice that the individuals from early medieval English sites are distinctly heterogeneous in the first two PCs and cover the full extent of the cline between the Bronze and Iron Age cluster and the early medieval cluster.

a, Present-day genomes from northwestern Europe. b, Published and novel ancient individuals in this study, projected onto a. Polygons indicate where two-thirds of the respective groups are located (England Bronze Age (BA) + Iron Age (IA) and North Sea IA + Early Middle Ages (EMA), respectively). The Scandinavian IA samples are connected with lines for clarity. For rough time boundaries of the samples used here, see Methods.
These genetic patterns suggest that early medieval individuals from England have variable amounts of CNE ancestry. Although most individuals from early medieval English sites cluster clearly with either present-day WBI samples or CNEs, many individuals fall between these two clusters, suggesting admixture between these ancestral groups. To quantitatively estimate these ancestry compositions, we decomposed ancestral sources using a supervised clustering approach implemented in the software ADMIXTURE48. Specifically, we assembled modern populations into two metapopulations that serve as proxies for the source ancestries in early medieval England defined above: CNE (n = 407) and WBI (n = 667). We confirmed that these two present-day metapopulations accurately represent the ancient admixture sources by testing their relationships to the ancient individuals from England using FST statistics and F4 statistics of the form F4(Yoruba, Test; WBI, CNE) (Extended Data Fig. 1 and 2). The resulting ancestry estimates for early medieval English individuals are indeed tightly congruent with both PCA PC1 position and F4 statistics (Pearson’s |r| > 0.9 between PCA, F4 and ADMIXTURE ancestry assessments).
Applying our CNE–WBI ancestry decomposition to prehistoric samples, we found the genome-wide CNE ancestry in Britain and Ireland to be very low before the Early Middle Ages (Extended Data Fig. 3). In Bell Beaker and Bronze Age individuals from England, CNE ancestry does not account for more than 1% (Fig. 3a). This cannot be explained by genetic drift due to the temporal gap between our present-day CNE proxy and the Bronze Age, as shown by F4 statistics (Extended Data Figs. 1 and 2), which are robust against such drift. Similar proportions were also measured during the Iron Age (1% on average). CNE ancestry increased only during the Roman period, to 15% on average, although this estimate is based on only seven individuals. Six of those seven Roman-era individuals are from a single site, Eboracum (present-day York); which was a Colonia, the highest rank of Roman city with a legionary fortress, and as such it may have attracted a more cosmopolitan population than most of the rest of England (Fig. 3b).

a, Individual supervised admixture results for Bronze Age (n = 140), Iron Age (n = 304) and Early Middle Age (n = 285) genomes from England. The symbols and colours of early medieval sites correspond to Fig. 1. b, Mean CNE and WBI ancestry estimates of British–Irish sites from the Bronze, Iron and Early Middle Ages.
In contrast to these previous periods, the majority of the early medieval individuals from England in our sample derive either all or a large fraction of their ancestry from continental northern Europe, with CNE ancestry of 76 ± 2% on average (Methods). Although CNE ancestry is predominant in central and eastern England, it is much less prevalent in the south and southwest of England, and absent in the one site that we analysed from Ireland (Fig. 3b). Moreover, we observed differences in continental ancestry not only between but also within sites. Although we estimate CNE ancestry to be prevalent across eastern English cemeteries, there was considerable variation at the individual level, ranging from 0% to 100% of CNE ancestry within a site. For example, at Hatherdene Close (n = 17) in Cambridgeshire, we estimated a mean CNE ancestry of approximately 70%, with eight individuals exhibiting exclusively CNE ancestry, but three individuals having low or zero CNE ancestry. Overall, these patterns of genetic heterogeneity, from the transregional to the family level, are consistent with continuous interaction between the Iron Age-derived Romano-British population and migrants from the continent.
We find no significant differences of CNE or WBI ancestry between male and female individuals (Supplementary Note 7), and find individuals of both ancestries within prominent and/or furnished burials. In England overall, individuals with CNE ancestry (here and in the following, CNE means more than 50% CNE, and WBI means less than 50% CNE) are more likely to be found with grave goods than individuals with WBI ancestry (Fisher’s exact test P = 0.016). Of note, this appears to be driven by female individuals with CNE ancestry who are more likely to be found with grave goods (P = 0.001), and in particular brooches (P = 0.012), than female individuals with WBI ancestry (both based on Fisher’s exact test). However, graves belonging to male individuals with CNE ancestry are just as likely to have grave goods (P = 0.57) or weapons (P = 1) as those with WBI ancestry (both based on Fisher’s exact test). This is underlined by specific examples, such as a near 100% WBI male burial in grave 37 at Updown Eastry found with a seax under a barrow marked by a ring ditch, indicating a prominent weapon burial associated with a prominent person or status (Supplementary Fig. 1.1).
This pattern is also visible in East Anglia specifically, where individuals with CNE ancestry more often have grave goods (P = 0.014). This is also significant when considering only female individuals (P = 0.025), but not when considering females with brooches, which display gender-related status (P = 0.197). At the site level, these patterns are partly significant at Hatherdene Close (P = 0.015, 0.036 and 0.1, respectively). Treating ancestry not as a binary but as a continuous variable largely agrees with the previous results (see Supplementary Note 7), with a notable exception of West Heslerton, which stands out from this overall pattern, where men with a greater proportion of CNE ancestry are more likely to be found with weapons (Wilcoxon rank sum P = 0.02, although non-significant with Fisher’s exact test P = 0.53), which is the only significant signal of this type that we found (Lakenheath also displays many CNE burials with weapons, but with limited sample size).
There are notable individual exceptions to these patterns, such as a predominantly (60%) WBI burial at grave 80 in Oakington, found with the skeleton of a cow, silvered disc brooches and a chatelaine, and interred under a barrow, which is one of the more notable or wealthy burials in this cemetery49 (Supplementary Fig. 1.4). We note that several burials with weapons that were previously identified as female and discussed in the literature23,50 have turned out to be genetically male in our analysis (see the highlighted entries in Supplementary Table 1). Of note, however, a single individual still displays a sex–gender difference: a teenage boy buried with an equal-arm brooch, beads and a knife (grave 122 in West Heslerton).
In Dover Buckland, one of the most comprehensively sampled cemeteries in our dataset, we observed the mixing of genetic and cultural identities at the family level. For example, we found a group of relatives, spanning at least three generations, who all exhibit unadmixed CNE ancestry (Extended Data Fig. 4a,c). Down the pedigree, we then see the integration of a female into this group, who herself had unadmixed WBI ancestry (grave 304), and two daughters (graves 290 and 426), consequently of mixed ancestry. WBI ancestry entered again one generation later, as visible in near 50:50 mixed-ancestry grandchildren (graves 414, 305 and 425). Grave goods, including brooches and weapons, are in fact found on both sides of this family tree, pre-mixing and post-mixing (for example, in the youngest and mixed generation, we found both weapons, beads and pin, and their mother with a brooch). Although the first mixed generation is buried in close proximity to each other, the grandchildren are elsewhere on the site, although placed together (Extended Data Fig. 4b).
A quite different pattern is observed at Apple Down, which is among the most western sites that we have analysed. Here graves can be classified into distinct burial configurations according to orientation, location and frequency of artefacts. We found that burials with CNE ancestry are more often buried in configuration A (located towards the middle of the site and with east–west burial orientation) than in configuration B (located more towards the edges and with north–south orientation)49 (Fisher’s exact test P = 0.048). This shows that there is a significant difference within the treatment of individuals according to their ancestry, a finding very similar to those at early medieval cemeteries in Hungary and Italy with respect to northern versus southern European ancestry51.
Ancestry sources across the North Sea
Our new continental medieval data from regions bordering the North Sea provide a unique opportunity to further investigate the potential source of the CNE-related ancestry increase that we have described above (Supplementary Note 3). To this end, we first selected individuals who, according to our CNE–WBI decomposition, are of unadmixed CNE ancestry (CNE of more than 95%; from here from as England EMA CNE). For each site in the continental dataset, we then tested whether its individuals were genetically similar to the England EMA CNE group (n = 109) in terms of allele frequencies. Among the continental medieval groups analysed, sites from both northern Germany and Denmark are indeed indistinguishable from England EMA CNE individuals (Fig. 4). Consistently, England EMA CNE and medieval individuals from Lower Saxony exhibit almost identical genetic affinities and ancestry components (Extended Data Fig. 5 and Supplementary Fig. 3.2), possess the highest level of genetic similarity (based on F2, F3, F4 and FST statistics) (Extended Data Fig. 5 and Supplementary Fig. 3.8) and are symmetrically related to most ancient and modern populations (Supplementary Table 3.12). Together, this suggests that they are likely derived from the same source population. Using the LOCATOR52 software, which uses machine learning to map individuals into geographical space based on their genetic profiles, we infer a region spanning the northern Netherlands to the southernmost tip of Sweden as a putative source for the England EMA CNE ancestors, with a large proportion of individuals being assigned to Lower Saxony (see Methods) (Fig. 4). This similarity adds to previous evidence from the material culture and burial practices, especially between the Elbe-Weser region and the early Anglo-Saxon cemeteries, from which the archaeological migration discourse initially arose53. However, we also note the strong genetic homogeneity among most analysed sites in the northern Netherlands, northern Germany and Denmark (Supplementary Note 4), implying that, during the Early Middle Ages, the continental North Sea and adjacent western Baltic Sea area was a genetic continuum spanning most of the western North European plain without major geographical substructure (Supplementary Fig. 4.1,4.4). This, together with genetic backflow from the British–Irish Isles into continental Europe (Supplementary Table 4.2 and Supplementary Fig. 4.2,4.4), reflects the inferred linguistic history54 and precludes further identification of specific microregions that contributed gene flow to Britain. We note that, although our screening of plausible medieval continental sites is broad, it could overemphasize later developments of the genetic structure due to the increased replacement of cremation burials by inhumations on the continent. It also has a specific caveat in Scandinavia, where our medieval reference populations are mostly from Viking-era burials, which have diverse and mixed ancestries that may not be representative of the earlier populations there42,44.

Shown are (1) continental sites that are genetically indistinguishable from the more than 95% CNE EMA English (England EMA CNE) population using qpWave and provide fitting P values as source in a two-way qpAdm model of England EMA, as well as (2) the predicted locations for 72 England EMA CNE genomes using LOCATOR52. The red dashed line marks where 95% of the qpAdm and qpWave data are located.
Already during the Early Middle Ages, several individuals from multiple sites exhibit modest degrees of excess affinity (5.4%) to present-day individuals from the Scandinavian peninsula (Supplementary Fig. 6.2a), indicating additional sources. Although close cultural contacts to the Scandinavian peninsula are attested in the archaeological record3, we did not find this genetic variation to be geographically stratified within early medieval England (Supplementary Fig. 6.2b). This Scandinavian Peninsula-related ancestry increases substantially (to 30.6%) only during the Viking period (Supplementary Note 6).
Having established the close relatedness between specific continental regions and the individuals from early medieval England, we modelled the latter more directly using ancient source populations with the method qpAdm35. Specifically, we pooled ancient individuals in England by site and modelled each group as being admixed between two sources: one represented by pooled Iron Age/Roman period individuals from England, and the other represented by pooled early medieval individuals from Lower Saxony (from here known as LowerSaxony EMA). The resulting admixture proportions obtained in this way for early medieval sites in England are strongly correlated with our mean estimates from supervised ADMIXTURE above, as well as site-wise F4 statistics and mean PCA position (Pearson’s |r| > 0.9 between all four ancestry assessments) (Extended Data Fig. 6).
Using this model, we detected an average of 86 ± 2% ancestry from Lower Saxony across all early medieval sites in England, only slightly higher than the 76 ± 2% estimated using present-day source populations and supervised ADMIXTURE. At a regional scale, we observed more ancestry from Lower Saxony in eastern England than in the southwest, consistent with ancestry arriving from the east, either in one event or over a continuous time period. Our estimate of genome-wide ancestry is supported by independent evidence for population turnover from uniparental markers (Supplementary Fig. 2.7). Before the Middle Ages, post-Neolithic individuals from Britain and Ireland carried overwhelmingly the major Y chromosomal haplogroup R1b-P312, especially the sub-haplogroup R-L21 (refs. 39,41), which in present day shows a cline across the region, with highest frequencies in the west55,56. By contrast, the early medieval population of England exhibits a substantial fraction of continental-derived haplotypes belonging to haplogroups R1b-U106, R1a-M420, I2a1-L460 and I1-M253, which are commonly found in northern and central Europe (and are also common among ancient continental individuals including the ones that we report). In particular, Y chromosomal haplogroups I1-M253 and R1a-M420 were absent from our Bronze, Iron and Roman Age British and Irish individuals, but were identified in more than one-third of our individuals from early medieval England. Overall, haplogroups absent in Bronze and Iron Age England represent at least 73 ± 4% of the Y chromosomes in our early medieval English sample, mirroring the turnover estimates from autosomal data. Similarly, mitochondrial genomes show evidence of female lineage population turnover from regions bordering the North Sea (Supplementary Note 2 and Supplementary Fig. 2.4).
Estimates of continental ancestry on the X chromosomes (Supplementary Fig. 2.11), as well as estimates of source origin of Y chromosomal haplogroups (Supplementary Fig. 2.16) point to no significant difference between male-specific lineages and autosomal admixture estimates (Supplementary Note 2). Although neither mitochondrial, Y chromosomal or X chromosomal data can exclude subtle levels of sex bias during the admixture (Supplementary Note 2), they are also consistent with a model of no sex bias, suggesting that the migrants included both men and women who mixed at similar levels with the local population. We note that absence of sex bias during the early medieval CNE–WBI admixture does not exclude possibilities for sex bias in the later admixture processes that caused the dilution of CNE ancestry in present-day England observed below.
Recent population shifts in England
Although the most prominent signal of admixture in early medieval England is the rise in ancestry related to medieval and modern continental northern Europe, we found that several English sites include genomes that could not be explained as products of admixture between the two hypothesized ancestral gene pools—England IA or LowerSaxony EMA—using qpAdm57. Instead, these genomes have additional continental western and southern European ancestry (Supplementary Note 5). This ancestry is genetically very similar to Iron Age genomes from France5,6 (France IA) (Extended Data Fig. 7, Supplementary Table 5.1 and Supplementary Fig. 5.3,5.4). The majority of this French Iron Age-derived ancestry is found in early medieval southeastern England, namely, at the sites of Apple Down, Eastry, Dover Buckland and Rookery Hill, where it constitutes up to 51% of the ancestry identified (Extended Data Fig. 8a and Supplementary Table 5.4).
The appearance of France IA-related ancestry in early medieval England anticipates a pattern that we also clearly see in the present-day English population structure, in which we found that the same two-way CNE–WBI model that fits most ancient English fails for the modern population (Supplementary Fig. 5.8,5.11). Indeed, the missing component in the modern English population appears to be represented well by France IA (Supplementary Table 5.2 and Supplementary Fig. 5.2).
Using qpAdm (Methods), most present-day Scottish, Welsh and Irish genomes can be modelled as receiving most or all of their ancestry from the British Bronze or Iron Age reference groups, with little or no continental contribution. By contrast, for all present-day English samples the simple two-way admixture model (England LIA + England EMA CNE) fails. By extending our model to a three-way with added France IA as a third component, we now obtain fitting models (Supplementary Fig. 5.11,5.21). We estimate that the ancestry of the present-day English ranges between 25% and 47% England EMA CNE-like, 11% and 57% England LIA-like and 14% and 43% France IA-like. There are substantial genetic differences between English regions (Fig. 5a), with less ancient continental ancestry (England EMA CNE or France IA related) evident in southwestern and northwestern England as well as along the Welsh borders (Fig. 5c). By contrast, we saw peaks in CNE-like ancestry of up to 47% for southeastern, eastern and central England, especially Sussex, the East Midlands and East Anglia. We found substantial France IA ancestry only in England, but not in Wales, Scotland or Ireland, following an east-to-west cline in Britain (Pearson’s |r| > 0.86), accounting for as much as 43% of the ancestry in East Anglia (Fig. 5d). Very similar results were produced using LowerSaxony EMA as a source for CNE ancestry (Extended Data Fig. 8b). One potential caveat in this analysis is our relatively sparse Roman sample from England, where we particularly lack samples from the south, which might have pre-existing France IA-related ancestry. We, therefore, turned to one of our early medieval sites, the post-Roman cemetery of Worth Matravers at the southern coast of Dorset, whose individuals have nearly no CNE ancestry (less than 6% on average), and thus may serve as a more temporally close proxy for post-Roman Britain before the arrival of CNEs. When used as a source in our model, we found that the estimates of France IA-related ancestry in present-day England changed by less than 3% on average across the regions (Fig. 5b), suggesting that France IA-related ancestry entered England to a substantial amount after the Roman period. We note that a model involving southern or western European-like ancestry in England has been previously proposed36 on the basis of present-day samples, but we can now go further and delineate this third component more clearly against the CNE-like immigrant gene pool making up the majority of the early medieval individuals from England that we studied.
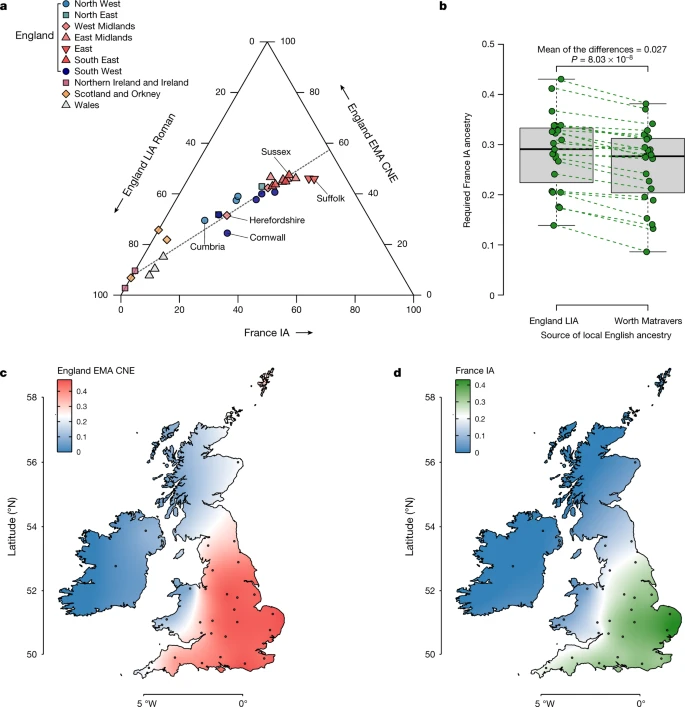
a, Ternary plot of present-day British–Irish populations as a three-way admixture between late Iron Age and Roman England (England LIA Roman) (n = 32), France IA (n = 26) and England EMA CNE (n = 109). b, Boxplot comparison of France IA ancestry proportions in 23 English PoBI sampling regions using either England LIA Roman (n = 32) or Worth Matravers (n = 16) as source for local British ancestry in qpAdm. The P value obtained from a two-sided paired Student’s t-test is shown. The bounds of the box represent the 25th and 75th percentile, the centre represents the median, and the whiskers represent the minimum and maximum values in the data. Dashed lines connect points from the same region. c, Geographical distribution of the England EMA CNE, ancestries based on the interpolation of 31 present-day population estimates. The coordinates of the sample collection districts approximate the centroids of the averaged birthplaces of the grandparents. d, Same as c, but for France IA.
Our three-way population model for present-day England supports a view of post-Roman English genetic history as punctuated by gene flow processes from at least two major sources: first, the attested arrival of CNE ancestry during the Early Middle Ages from northern Germany, the Netherlands and Denmark, and second, the arrival of ancestry related to France IA. Although we cannot precisely date the order of those arrivals, at least substantial amounts of France IA-related ancestry seem to be absent in northern and eastern England during the Early Middle Ages and therefore must have arrived there subsequently. In other parts of England, however, it may have entered together with CNE ancestry or even earlier. Notably in southern England, namely, Eastry, Apple Down and Rookery Hill, several early medieval individuals already exhibit France IA-related ancestry, which probably results, at least in part, from localized mobility between the south of England and the Frankish areas of Europe during the Early Middle Ages (Extended Data Fig. 8a). Indeed, Frankish material culture is evident in these regions, particularly in Kent and Sussex58,59,60. Admixture from this second source is, therefore, unlikely to have resulted from a single discrete wave. More plausibly, it resulted from pulses of immigration or continuous gene flow between eastern England and its neighbouring regions.
Discussion
The ‘Anglo-Saxon settlement’ is among the most intensely debated topics in British history, but much of the discussion remains anchored to the contents of Bede’s Ecclesiastical History and the Anglo-Saxon Chronicle18. These early writings defined the settlement as a single event, or a series of events, tied to the immediate aftermath of the Roman administration in the fifth to sixth century. In the archaeological and historical debate, this has been described as happening to varying degrees; as the Adventus Saxonum (a folk migration of named Germanic tribes), an invasion or the movement of a limited number of elite male migrants18,61. To this day, little agreement has been reached over the scale of migration, the mode of interaction between locals and newcomers, or how the transformation of the social, material, and linguistic or religious spheres was achieved. Here we provide strong evidence of large-scale early medieval migration across the North Sea zone and extend its temporal scope. In particular, we show that these migrations started earlier than previously assumed, as evidenced by individuals with CNE ancestry from later Roman contexts, and continued throughout the middle Anglo-Saxon period. Our results from middle Saxon sites such as Sedgeford push the estimated dates of arrival of CNE ancestry to as late as the eight century and merge these events with interpersonal mobility from Sweden and other Scandinavian regions during the later Viking invasion and settlement. Together, these migrations appear to be part of a continuous movement of people from across the North Sea to Britain from the later Roman period into the eleventh century ce.
Our results overwhelmingly support the view that the formation of early medieval society in England was not simply the result of a small elite migration18,61, but that mass migration from afar must also have had a substantial role. We identified numerous individuals with only continental ancestry, suggesting that many of them were migrants themselves or were their unadmixed descendants. Both the lack of genetic evidence for male sex bias, and the correlation between ancestry and archaeological features, point to women being an important factor in this migration. Although men with migrant and local ancestry were buried in similar ways, women with migrant ancestries were more often found with grave goods than women with local ancestry. This could point to social stratification, or plausibly might simply reflect the degree to which women of local ancestry were integrated into the emerging CNE families. It is clear, however, that these social differences are subtle, given that we did not find evidence for this pattern in male burials, and that we found significant regional and site-level differences. Previous hypotheses about the social mechanisms in this migration have included partial social segregation62, elite migration18,61, substantial population replacement34 or no migration at all1,22. Our combined genetic and archaeological analysis point to a complex, regionally contingent migration with partial integration that was probably dependent on the fortunes of specific families and their individual members.
In present-day Britain, we saw substantial northern continental ancestry, albeit at a lower level than during the early medieval period, pointing to a lasting demographic impact of the ‘Anglo-Saxon’ migrations. Specifically, in early medieval western England, Wales and Scotland, and more generally in England during the Norman period, further aDNA sampling may clarify how CNE ancestry spread and was subsequently diluted. Beyond the substantial early medieval immigration of northwestern continental European people found here, we have also identified a second major source of continental ancestry in modern Britain from sources more to the European south and west. This second ancestry component is already evident in our early medieval samples. In Southeast England specifically, individuals at several sites show ancestry whose closest match is in modern-day western Germany, Belgium and/or France, which matches the Frankish connections seen in the archaeological record for these regions. Our data and analyses indicate that this second genetic introgression continued further into the Middle Ages and potentially beyond.
Methods
Study design
Archaeological research
Provenance information for samples from all archaeological sites are given in Supplementary Information Section 1, together with short descriptions of each site, the institution owning the samples (or custodians of the samples), the responsible coauthor who obtained permission to analyse and the year of the permission granted.
Sampling
Sampling of 494 bone and teeth samples took place in clean-room facilities dedicated to aDNA work, for 296 samples at the Max Planck Institute for Science of Human History in Jena (MPI-SHH), for 65 at the Department of Biological and Geographical Sciences at the University of Huddersfield, for 33 at the Department of Genetics, Harvard Medical School (HMS), for 32 at the Institute for Scientific Archaeology of the Eberhard Karls University Tübingen, for 31 at the Leiden University Medical Centre in Leiden, for 15 at the Globe Institute of the University of Copenhagen, for 12 at the Australian Centre for Ancient DNA at the University of Adelaide, and for 10 at the Research Laboratory for Archaeology, University of Oxford. The sampling workflow included documenting and photographing the provided samples. For teeth processed at the MPI-SHH, we cut along the cementum–enamel junction and collected powder by drilling into the pulp chamber. The teeth processed at the Leiden University Medical Centre were sampled according to a previously published paper63. For the petrous bones, we either cut the petrous pyramid longitudinally to drill the dense part directly from either side64 or applied the cranial base drilling method as previously described65. We collected between 30 and 200 mg of bone or tooth powder per sample for DNA extractions.
DNA extraction
At MPI-SHH and HMS, aDNA was extracted following a modified protocol66, as described in www.protocols.io/view/ancient-dna-extraction-from-skeletal-material-baksicwe, in which we replaced the extended-MinElute-column assembly for manual extractions with columns from the Roche High Pure Viral Nucleic Acid Large Volume Kit67, and for automated extraction with a protocol that replaced spin columns with silica beads in the purification step68. Extraction of aDNA at the Leiden University Medical Centre, at the Globe Institute and at the University of Adelaide followed the protocols of Kootker et al.63, Damgaard et al.69 and Brotherton et al.70, respectively. Extraction of aDNA at the Universities of Oxford and Huddersfield followed a published protocol71.
Library construction
We generated 104 double-indexed72 double-stranded libraries using 25 µl of DNA extract and following established protocols73. We applied the partial UDG (UDG half)74 protocol to remove most of the aDNA damage while preserving the characteristic damage pattern in the terminal nucleotides. For 375 extracts, we generated double-indexed single-stranded libraries75 using 20 µl of DNA extract and applied no UDG treatment.
Shotgun screening, capture and sequencing
Libraries produced at MPI-SHH were sequenced in-house on an Illumina HiSeq 4000 platform to an average depth of 5 million reads and after demultiplexing processed through EAGER76. After an initial quality filter based on the presence of aDNA damage and endogenous DNA higher than 0.1%, we subsequently enriched 439 libraries using in-solution capture probes synthesized by Agilent Technologies for approximately 1240K SNPs along the nuclear genome77. The captured libraries were sequenced for 20–40 million reads on average using either a single end (1 × 75 bp reads) or paired-end configuration (2 × 50 bp reads). In addition, 40 genomes were shotgun sequenced for 225 million reads on average to low coverage. For the 120 samples processed at HMS, we enriched for sequences overlapping approximately 1240K SNPs77 as well as the mitochondrial genome78, and sequenced on Illumina NextSeq500 instruments for 2 × 76 cycles, or on HiseqX10 instruments for 2 × 101 cycles (reading out both indices) approximately until the point in which every additional 100 sequences generated yielded fewer than one additional SNP with data.
aDNA data processing
Read processing and aDNA damage
For data produced at the MPI-SHH, after demultiplexing based on a unique pair of indexes, raw sequence data were processed using EAGER76. This included clipping sequencing adaptors from reads with AdapterRemoval (v2.3.1)79 and mapping of reads with Burrows–Wheeler Aligner (BWA)80 v0.7.12 against the human reference genome hg19, with seed length (-l) disabled, maximum number of differences (-n) of 0.01 and a quality filter (-q) of 30. We removed duplicate reads with the same orientation and start and end positions using DeDup76 v0.12.2. Terminal base deamination damage calculation was done using mapDamage81 v2.0.6, specifying a length (-l) of 100 bp. For the 107 libraries that underwent UDG half treatment, we used BamUtil v1.0.14 (https://genome.sph.umich.edu/wiki/BamUtil:_trimBam) to clip two bases at the start and end of all reads for each sample to remove residual deaminations, thus removing genotyping errors that could arise due to aDNA damage.
For data produced at the HMS, after trimming barcodes and adapters57, we merged read pairs with at least 15 bp of overlap, allowing no more than one mismatch if base quality was at least 20, or up to three mismatches if base qualities were less than 20. We chose the nucleotide of the higher quality in case of a conflict while setting the local base quality to the minimum of the two using a custom toolkit (https://github.com/DReichLab/ADNA-Tools). We aligned merged sequences to human genome hg19 using BWA80 v0.7.15 with a maximum number of differences (-n) of 0.01, a maximum number of gap opens (-o) of 2 and seed length (-l) of 16,500. PCR duplicates were identified by tagging all aligned sequences with the same start and stop positions and orientation and, in some cases, in-line barcodes using Picard MarkDuplicates (http://broadinstitute.Github.io/picard/). We only considered sequences that spanned at least 30 bp, and subsequently selected a single copy of each such sequence that had the highest base-quality score. To remove aDNA damage, we trimmed the last two bases of each sequence for UDG-treated libraries and the last five for non-UDG-treated libraries.
Sex determination
To determine the genetic sex of ancient individuals processed at the MPI-SHH, we calculated the coverage on the autosomes as well as on each sex chromosome and subsequently normalized the X reads and Y reads by the autosomal coverage82. For that, we used a custom script (https://github.com/TCLamnidis/Sex.DetERRmine) for the calculation of each relative coverage as well as their associated error bars83. Female individuals were expected to have an X rate of 1 and a Y rate of 0, whereas male individuals were expected to have a rate of 0.5 for both X and Y chromosomes. For individuals processed at the HMS, we calculated the ratio of sequences mapping to the Y chromosome to the sum of sequences mapping to the X and Y chromosome for the 1240K data. A ratio less than 3% is consistent with a female individual and a ratio higher than 35% is consistent with a male individual6.
Contamination estimation
We used the Analysis of Next Generation Sequencing Data (ANGSD) package84 (v0.923) to test for heterozygosity of polymorphic sites on the X chromosome in male individuals, applying a contamination threshold of 5% at the results of method two. For male and female samples, we estimated contamination levels on the mitochondrial DNA either using Schmutzi85 (v1.5.4) by comparing the consensus mitogenome of the ancient sample to a panel of 197 worldwide mitogenomes as a potential contamination source (MPI-SHH), or by estimating the match rate to the consensus sequence using contamMix86 v1.0-12 (HMS), applying a contamination threshold of 5%. We used PMDtools87 (v0.50) to isolate sequences from each sample that had clear evidence of contamination (over 5% on the X chromosome or mitogenome) according to the post-mortem damage score (PMD score > 3, using only bases with phred-scaled quality of at least 30 to compute the score), and performed contamination estimation again. If a sample scored below the threshold, it was included in the analysis and modelling. If the authenticity of a sample could not be verified or falsified, it was included in population genetic analyses but not used for modelling. In summary, the median mitochondrial DNA contamination is 1.0%, and the median X chromosome contamination is 1.1% (after PMD filtering).
Genotyping
We used the program pileupCaller (v1.4.0.2) (https://github.com/stschiff/sequenceTools.git) to genotype the trimmed BAM files of UDG half libraries. A pileup file was generated using samtools mpileup with parameters -q 30 -Q 30 -B containing only sites overlapping with our capture panel. From this file, for each individual and each SNP on the 1240K panel57,88,89, one read covering the SNP was drawn at random, and a pseudohaploid call was made, that is, the ancient individual was assumed homozygous for the allele on the randomly drawn read for the SNP in question. For libraries that underwent no UDG treatment, we used the parameter -SingleStrandMode, which causes pileupCaller to ignore reads aligning to the forward strand at C/T polymorphisms and at G/A polymorphisms to ignore reads aligning to the reverse strand, which should remove post-mortem damage in aDNA libraries prepared with the non-UDG single-stranded protocol.
Mitochondrial and Y chromosome haplogroup assignment
To process mitochondrial DNA data generated at the MPI-SHH, we extracted reads from 1240K data using samtools90 v1.3.1 and mapped these to the revised Cambridge reference sequence. At the HMS, we aligned merged sequences to the mitochondrial genome RSRS91. We subsequently called consensus sequences using Geneious92 R9.8.1 and used HaploGrep 2 (ref. 93) v2.4.0 (https://haplogrep.uibk.ac.at/; with PhyloTree version 17-FU1) to determine mitochondrial haplotypes. For the male individuals processed at the MPI-SHH, we used pileup from the Rsamtools package to call the Y chromosome SNPs of the 1240K SNP panel (mapping quality of 30 or more and base quality of 30 or more). We then manually assigned Y chromosome haplogroups using pileups of Y SNPs included in the 1240K panel that overlap with SNPs included on the ISOGG SNP index v.15.73 (Y-DNA Haplogroup Tree 2019–2020; 2020.07.11). For male individuals processed at the HMS, we automatically determined Y chromosome haplogroups using both targeted SNPs and off-target sequences aligning to the Y chromosome based on comparisons to the Y chromosome phylogenetic tree from Yfull version 8.09 (https://www.yfull.com/)6.
Kinship estimation
We calculated the PWMR94 in all pairs of individuals from our pseudo-haploid dataset to double check for potential duplicate individuals and to determine first-degree, second-degree and third-degree relatives. For this purpose, we also used READ95 to determine first-degree, second-degree and third-degree relatedness among individuals based on the proportion of non-matching alleles (P0) in nonoverlapping windows of 1 Mb and to calculate standard errors. We also used the method LcMLkin96, which uses genotype likelihoods to estimate the three k-coefficients (k0, k1 or k2), which define the probability that two individuals have zero, one or two alleles identical by descendent at a random site in the genome. We performed LcMLkin to distinguish between possible parent–offspring or sibling relationships.
Population genetic analysis
Dataset
We merged our aDNA data with previously published datasets of 4,336 ancient individuals reported by the Reich laboratory in the Allen Ancient DNA Resource v.50.0 (https://reich.hms.harvard.edu/allen-ancient-dna-resource-aadr-downloadable-genotypes-present-day-and-ancient-dna-data). We assembled a dataset from mostly European populations for genome-wide analyses36,45,46,47,97,98,99,100,101,102,103. This modern set includes 10,176 individuals (Supplementary Note 2). Loci and individuals with less than 95% call rate as well as a 15-Mb region surrounding the HLA region36 were removed and loci on three previously reported long-range linkage disequilibrium regions on chromosomes 6, 8 and 11 (refs. 104,105) were pruned using PLINK106 (v1.90b3.29). aDNA data were merged to this dataset, correcting for reference allele and strand flips. We kept 445,171 autosomal SNPs after intersecting autosomal SNPs in the 1240K capture with the modern analysis set.
Abbreviations
We used the following abbreviations in population labels: N, Neolithic; C, Chalcolithic; EBA, Early Bronze Age; MBA, Middle Bronze Age; LBA, Late Bronze Age; Iron Age, IA; RA, Roman Age; EMA, Early Middle Ages; MA, Middle Ages. In Britain, these periods roughly correspond to the following simplified time ranges: Neolithic: 4000–2500 bce, Chalcolithic and EBA: 2500–1600 bce; MBA: 1600–1200 bce; LBA: 1200–800 bce; IA: 800 bce to 400 ce; EMA 400–1000 ce.
PCA
We carried out PCA using the smartpca software v16000 from the EIGENSOFT package (v6.0.1)107. We computed PCs on three different sets of modern European populations (Supplementary Note 2) and projected ancient individuals using lsqproject: YES.
F statistics
F3 and F4 statistics were computed with ADMIXTOOLS108 v3.0 (https://github.com/DReichLab). F3 statistics were calculated using qp3Pop (v435). For F4 statistics, we used the qpDstat (v755) and with the activated F4 mode. Significant deviation from zero can be interpreted as rejection of the tree population typology ((outgroup, X);(Pop1, Pop2)). Under the assumption that no gene flow occurred between Pop1 and Pop2 and the outgroup, a positive f-statistic suggests affinity between X and Pop2, whereas a negative value indicates affinity between X and Pop1. Standard errors were calculated with the default block jackknife 5 cM in size.
Fixation index
We calculated FST using smartpca software v16000 from the EIGENSOFT package (v6.0.1)107 with the fstonly, inbreed and fsthiprecision options set to YES.
Maximum likelihood tree
We constructed maximum likelihood trees using TreeMix (v1.12)109. For each tree, we performed a round of global rearrangements after adding all populations (-global) and calculated 100 bootstrap replicates to assess the uncertainty of the fitted model (-bootstrap). Sample size correction was disabled.
Inference of mixture proportions
We estimated ancestry proportions using qpWave57,110 (v410) and qpAdm57 (v810) from ADMIXTOOLS108 v3.0 with the allsnps: YES option and a basic set of 11 outgroups: YRI.SG, Poland, Finland, Sweden, Denmark, Ireland, Wales, Italy, Spain, Belgium and the Netherlands. For some analyses, we added additional outgroups to this basic set (Supplementary Notes 4–6).
Prediction of geographical origins
LOCATOR52 (v1.2) was run using a geolocated reference panel consisting of 670 Bronze Age, Iron Age and medieval European samples with 1X coverage higher than 50% and considering only polymorphisms covered at least in 50% of the samples, leaving a total of 920,060 SNPs. Default parameters were used, except that the width of each neural layer was 512 and -imputed-missing was set to YES. The best run was selected as the one showing the lowest validation error and the highest R2 numbers from a total of 40 independent runs.
ADMIXTURE analysis
We performed model-based clustering analysis using ADMIXTURE48 (v1.3). We used ADMIXTURE in supervised mode, in which we estimated admixture proportions for the ancient individuals using modern reference populations at various K values (Supplementary Notes 3–6). These analyses were run on haploid data with the parameter –haploid set to all (="*"). Standard errors for point estimates were calculated using 1,000 bootstrap replicates with the -B parameter. To obtain point estimates for populations, we averaged individual point estimates and calculated the standard error of the mean (s.e.m.): s.e.m. = \(\frac{\sigma }{\surd n}\). We found that this better reflects the diversity within the population than a propagation of error approach, which underestimates the variance within the point estimate sample. For unsupervised ADMIXTURE analysis (Supplementary Notes 3 and 5), we carried out linkage disequilibrium pruning on the dataset using PLINK106 with the flag–indep-pairwise 200 25 0.4, leaving 306,393 SNPs. We ran ADMIXTURE with the cross-validation (–cv.) flag specifying from K = 2 to K = 10 clusters, with five replicates for each value of K. For each value of K, the replicate with highest log likelihood was kept.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Raw sequence data (bam files) from the 479 newly reported ancient individuals will be available before publication from the European Nucleotide Archive under accession number PRJEB54899. Published genotype data for the present-day British sample are available from the WTCCC via the European Genotype Archive (https://www.ebi.ac.uk/ega/) under accession number EGAD00010000634. Published genotype data for the present-day Irish sample are available from the WTCCC via the European Genotype Archive under accession number EGAD00010000124. Published genotype data for the rest of the present-day European samples are available from the WTCCC via the European Genotype Archive under accession number EGAD00000000120. Published genotype data for the Dutch samples are available by the GoNL request process from The Genome of the Netherlands Data Access Committee (DAC) (https://www.nlgenome.nl). The Genome Reference Consortium Human Build 37 (GRCh37) is available via the National Center for Biotechnology Information under accession number PRJNA31257. The revised Cambridge reference sequence is available via the National Center for Biotechnology Information under NCBI Reference Sequence NC_012920.1. Previous published genotype data for ancient individuals were reported by the Reich laboratory in the Allen Ancient DNA Resource v.50.0 (https://reich.hms.harvard.edu/allen-ancient-dna-resource-aadr-downloadable-genotypes-present-day-and-ancient-dna-data).
Code availability
All software used in this work is publicly available. Corresponding publications are cited in the main text and Supplementary information. List of software and respective versions: AdapterRemoval (v2.3.1), Burrows–Wheeler Aligner (v0.7.12), DeDup (v0.12.2), mapDamage (v2.0.6), BamUtil (v1.0.14), EAGER (v1), Sex.DetERRmine (https://github.com/TCLamnidis/Sex.DetERRmine) (v1.1.2), ANGSD (v0.915), Schmutzi (v1.5.4), contamMix (v1.0-12), PMDtools (v0.50), pileupCaller (v1.4.0.2), samtools (v1.3.1), Geneious R9.8.1, HaploGrep 2 (v2.4.0), READ (https://bitbucket.org/tguenther/read) (vf541d55), lcMLkin (https://github.com/COMBINE-lab/maximum-likelihood-relatedness-estimation) (v0.5.0), PLINK (v1.90b3.29), Picard tools (v2.27.3), ADNA-Tools (https://github.com/DReichLab/ADNA-Tools) (v3b4357d), smartpca (v16000; EIGENSOFT v6.0.1), qp3Pop (v.435; ADMIXTOOLS v3.0), qpDstat (v.755; ADMIXTOOLS v3.0), Treemix (v1.12), qpWave (v410), qpAdm (v.810), LOCATOR (v1.2) and ADMIXTURE (v1.3). The code used in Supplementary Note 2 (‘Estimating sex-biased ancestry from uniparental markers in the presence of variable admixture proportions’) can be found at https://github.com/stschiff/AngloSaxon_Y-chrom_sex-bias. Data visualization and descriptive statistical tests were performed in R (v4.1.1). The following R packages were used: Rsamtools (v2.12.0), binom (v1.1-1.1), ape (v.5.6-2), phytools (v1.0-3), psych (v2.2.5), vegan (v2.6-2), factoextra (v1.0.7), ggplot2 (v3.3.6), ggExtra (v0.10.0), ggforce (v0.3.3), rnaturalearth (v0.1.0), sf (v1.0.−8), raster (v3.5-21), elevatr (v0.4.2), rgdal (v1.5-32), spatstat (v2.3-4), maptools (v1.1-4), gstat (v2.0-9), sp (v1.5-0), labdsv (v2.0-1), igraph (v1.3.4), magrittr (v2.0.3), dplyr (v1.0.9), reshape 2 (v1.4.4) and tidyverse (v.1.3.2). Y chromosomal and mitochondrial DNA haplogroups were determined using the ISOGG SNP index (v15.73) and PhyloTree (v17-FU1) reference databases, respectively.
Change history
17 October 2022
A Correction to this paper has been published: https://doi.org/10.1038/s41586-022-05429-y
References
Fleming, R. The Material Fall of Roman Britain, 300–525 CE (Univ. Pennsylvania Press, 2021).
Hills, C. M. Did the people of Spong Hill come from Schleswig-Holstein? Studien zur Sachsenforschung 11, 145–154 (1999).
Hines, J. The Scandinavian Character of Anglian England in the Pre-Viking Period (Univ. Oxford, 1983).
Hines, J. The becoming of the English: identity, material culture and Language in early Anglo-Saxon England. Anglo Saxon Stud. Archaeol. Hist. 7, 49–59 (1994).
Brunel, S. et al. Ancient genomes from present-day France unveil 7,000 years of its demographic history. Proc. Natl Acad. Sci. USA 117, 12791–12798 (2020).
Patterson, N. et al. Large-scale migration into Britain during the Middle to Late Bronze Age. Nature 601, 588–594 (2021).
Hills, C. M. & Lucy, L. Spong Hill Part IX: Chronology and Synthesis (McDonald Institute for Archaeological Research, 2013).
Bruns, D. Germanic Equal Arm Brooches of the Migration Period Vol. 1,113 (Archaeopress, 2003).
Suzuki, S. The Quoit Brooch Style and Anglo-Saxon Settlement: a Casting and Recasting of Cultural Identity Symbols (Boydell Press, 2000).
Hamerow, H. Early Medieval Settlements: The Archaeology of Rural Communities in North-West Europe 400–900 (Oxford Univ. Press, 2020).
Hamerow, H. in Migrations and Invasions in Archaeological Explanation Vol. 664 (eds Chapman, J. & Hamerow, H.) 33–44 (Archaeopress, 1997).
Martin, T. F. The Cruciform Brooch and Anglo-Saxon England (Boydell & Brewer, 2015).
Hines, J. Clasps, Hektespenner, Agraffen: Anglo-Scandinavian Clasps of classes A–C of the 3rd to 6th Centuries A.D.: Typology, Diffusion and Function (Almqvist & Wiksell International, 1993).
Bruce-Mitford, R. A Corpus of Late Celtic Hanging-Bowls with an Account of the Bowls Found in Scandinavia (Oxford Univ. Press, 2005).
Scull, C. Further evidence from East Anglia for enamelling on Early Anglo-Saxon metalwork. Anglo Saxon Stud. Archaeol. Hist. 4, 117–122 (1985).
Gelling, M. in Anglo-Saxon Settlements (ed. Hook, D.) 59–76 (Blackwell, 1988).
Gelling, M. Signposts to the Past: Place-names and the History of England (Dent, 1978).
Lucy, S. The Early Anglo-Saxon Cemeteries of East Yorkshire: an Analysis and Reinterpretation (BAR Publishing, 2019).
Leeds, E. T. The archaeology of the Anglo-Saxon settlements. Nature 92, 369–369 (1913).
Myres, J. N. L. The English Settlements (Clarendon Press, 1968).
Kruse, P. Jutes in Kent? On the Jutish nature of Kent, southern Hampshire and the Isle of Wight. Probleme der Küstenforschung im südlichen Nordseegebiet 31, 243–376 (2007).
Prior, F. Britain AD: a Quest for Arthur, England and the Anglo-Saxons (HarperCollins, 2004).
Montgomery, J., Evans, J. A., Powlesland, D. & Roberts, C. A. Continuity or colonization in Anglo-Saxon England? Isotope evidence for mobility, subsistence practice, and status at West Heslerton. Am. J. Phys. Anthropol. 126, 123–138 (2005).
Budd, P., Millard, A., Chenery, C., Lucy, S. & Roberts, C. Investigating population movement by stable isotope analysis: a report from Britain. Antiquity 78, 127–141 (2004).
Hughes, S. S. et al. Anglo-Saxon origins investigated by isotopic analysis of burials from Berinsfield, Oxfordshire, UK. J. Archaeol. Sci. 42, 81–92 (2014).
Schiffels, S. et al. Iron Age and Anglo-Saxon genomes from East England reveal British migration history. Nat. Commun. 7, 10408 (2016).
Scull, C. in Europe Between Late Antiquity and the Middle Ages: Recent Archaeological and Historical Research in Western and Southern Europe Vol. 617 (eds Bintliffe, J. & Hamerow, H.) 71–83 (British Archaeological Reports, 1995).
Ulmschneider, K. in The Oxford Handbook of Anglo-Saxon Archaeology (eds Hamerow, H., Hinton, D. A. & Crawford, S.) 156–171 (Oxford Univ. Press, 2011).
Ward-Perkins, B. Why Did the Anglo-Saxons not become more British? Engl. Hist. Rev. 115, 513–533 (2000).
Coates, R. in Britons in Anglo-Saxon England (ed. Higham, N. J.) 172–191 (Boydell & Brewer, 2007).
Tristram, H. in Britons in Anglo-Saxon England (ed. Higham, N. J.) 192–214 (Boydell & Brewer, 2007).
Schrijver, P. in Language Contact and the Origins of the Germanic Languages Vol. 13 (ed. Schrijver, P.) 12–93 (Routledge, 2014).
Richards, M., Smalley, K., Sykes, B. & Hedges, R. Archaeology and genetics: analysing DNA from skeletal remains. World Archaeol. 25, 18–28 (1993).
Weale, M. E., Weiss, D. A., Jager, R. F., Bradman, N. & Thomas, M. G. Y chromosome evidence for Anglo-Saxon mass migration. Mol. Biol. Evol. 19, 1008–1021 (2002).
Capelli, C. et al. A Y chromosome census of the British Isles. Curr. Biol. 13, 979–984 (2003).
Leslie, S. et al. The fine-scale genetic structure of the British population. Nature 519, 309–314 (2015).
Martiniano, R. et al. Genomic signals of migration and continuity in Britain before the Anglo-Saxons. Nat. Commun. 7, 10326 (2016).
Allentoft, M. E. et al. Population genomics of Bronze Age Eurasia. Nature 522, 167–172 (2015).
Cassidy, L. M. et al. Neolithic and Bronze Age migration to Ireland and establishment of the insular Atlantic genome. Proc. Natl Acad. Sci. USA 113, 368–373 (2016).
Veeramah, K. R. et al. Population genomic analysis of elongated skulls reveals extensive female-biased immigration in Early Medieval Bavaria. Proc. Natl Acad. Sci. USA 115, 3494–3499 (2018).
Olalde, I. et al. The Beaker phenomenon and the genomic transformation of northwest Europe. Nature 555, 190–196 (2018).
Krzewińska, M. et al. Genomic and strontium isotope variation reveal immigration patterns in a Viking Age town. Curr. Biol. 28, 2730–2738.e10 (2018).
O’Sullivan, N. et al. Ancient genome-wide analyses infer kinship structure in an Early Medieval Alemannic graveyard. Sci. Adv. 4, eaao1262 (2018).
Margaryan, A. et al. Population genomics of the Viking world. Nature 585, 390–396 (2020).
International Multiple Sclerosis Genetics Consortium et al. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 476, 214–219 (2011).
Genome of the Netherlands Consortium. Whole-genome sequence variation, population structure and demographic history of the Dutch population. Nat. Genet. 46, 818–825 (2014).
Genetic Analysis of Psoriasis Consortium & The Wellcome Trust Case Control Consortium 2 et al. A genome-wide association study identifies new psoriasis susceptibility loci and an interaction between HLA-C and ERAP1. Nat. Genet. 42, 985–990 (2010).
Alexander, D. H., Novembre, J. & Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 19, 1655–1664 (2009).
Sayer, D. Early Anglo-Saxon Cemeteries (Manchester Univ. Press, 2020).
Lucy, S. in Invisible People and Processes: Writing Gender and Childhood into European Archaeology (eds Moore, J. S. E. & Scott, E.) 150–168 (Leicester Univ. Press, 1997).
Amorim, C. E. G. et al. Understanding 6th-century barbarian social organization and migration through paleogenomics. Nat. Commun. 9, 3547 (2018).
Battey, C. J., Ralph, P. L. & Kern, A. D. Predicting geographic location from genetic variation with deep neural networks. eLife 9, e54507 (2020).
Härke, H. Anglo-Saxon immigration and ethnogenesis. Mediev. Archaeol. 55, 1–28 (2011).
Hines, J. in Friesische Studien 2 Vol. 12 (eds Faltings, F. V., Walker, A. G. H. & Wilts, O.) 35–62 (Routledge, 1995).
Myres, N. M. et al. A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe. Eur. J. Hum. Genet. 19, 95–101 (2010).
Busby, G. B. J. et al. The peopling of Europe and the cautionary tale of Y chromosome lineage R-M269. Proc. R. Soc. B 279, 884–892 (2012).
Haak, W. et al. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 522, 207–211 (2015).
Brugmann, B. in The Pace of Change: Studies in Early-Medieval Chronology (eds Hines, J., Høilund Nielsen, K. & Siegmund, F.) 37–64 (Oxbow, 1999).
Soulat, J. in Studies in Early Anglo-Saxon Art and Archaeology: Papers in Honour of Martin Welch (eds Brookes, S., Harrington, S. & Reynolds, A.) 62–71 (British Archaeology Reports, 2011).
Evison, V. I. The Fifth-Century Invasions South of the Thames (Athlone Press, 1965).
Higham, N. J. Rome, Britain and the Anglo-Saxons (Seaby, 1992).
Thomas, M. G., Stumpf, M. P. H. & Härke, H. Evidence for an apartheid-like social structure in early Anglo-Saxon England. Proc. Biol. Sci. 273, 2651–2657 (2006).
Kootker, L. M. et al. Beyond isolation: understanding past human-population variability in the Dutch town of Oldenzaal through the origin of its inhabitants and its infrastructural connections. Archaeol. Anthropol. Sci. 11, 755–775 (2019).
Pinhasi, R. et al. Optimal ancient DNA yields from the inner ear part of the human petrous bone. PLoS ONE 10, e0129102 (2015).
Sirak, K. A. et al. A minimally-invasive method for sampling human petrous bones from the cranial base for ancient DNA analysis. Biotechniques 62, 283–289 (2017).
Dabney, J. et al. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl Acad. Sci. USA 110, 15758–15763 (2013).
Korlević, P. et al. Reducing microbial and human contamination in DNA extractions from ancient bones and teeth. Biotechniques 59, 87–93 (2015).
Rohland, N., Glocke, I., Aximu-Petri, A. & Meyer, M. Extraction of highly degraded DNA from ancient bones, teeth and sediments for high-throughput sequencing. Nat. Protoc. 13, 2447–2461 (2018).
Damgaard, P. B. et al. Improving access to endogenous DNA in ancient bones and teeth. Sci. Rep. 5, 11184 (2015).
Brotherton, P. et al. Neolithic mitochondrial haplogroup H genomes and the genetic origins of Europeans. Nat. Commun. 4, 1764 (2013).
Dulias, K. et al. Ancient DNA at the edge of the world: continental immigration and the persistence of Neolithic male lineages in Bronze Age Orkney. Proc. Natl Acad. Sci. USA 119, e2108001119 (2022).
Kircher, M., Sawyer, S. & Meyer, M. Double indexing overcomes inaccuracies in multiplex sequencing on the Illumina platform. Nucleic Acids Res. 40, e3 (2012).
Meyer, M. & Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, pdb.prot5448 (2010).
Rohland, N., Harney, E., Mallick, S., Nordenfelt, S. & Reich, D. Partial uracil-DNA-glycosylase treatment for screening of ancient DNA. Phil. Trans. R. Soc. B 370, 20130624 (2015).
Gansauge, M.-T. & Meyer, M. Single-stranded DNA library preparation for the sequencing of ancient or damaged DNA. Nat. Protoc. 8, 737–748 (2013).
Peltzer, A. et al. EAGER: efficient ancient genome reconstruction. Genome Biol. 17, 60 (2016).
Fu, Q. et al. An early modern human from Romania with a recent Neanderthal ancestor. Nature 524, 216–219 (2015).
Fu, Q. et al. DNA analysis of an early modern human from Tianyuan Cave, China. Proc. Natl Acad. Sci. USA 110, 2223–2227 (2013).
Schubert, M., Lindgreen, S. & Orlando, L. AdapterRemoval v2: rapid adapter trimming, identification, and read merging. BMC Res. Notes 9, 88 (2016).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Jónsson, H., Ginolhac, A., Schubert, M., Johnson, P. L. F. & Orlando, L. mapDamage2.0: fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 29, 1682–1684 (2013).
Mittnik, A., Wang, C.-C., Svoboda, J. & Krause, J. A molecular approach to the sexing of the triple burial at the Upper Paleolithic site of Dolní Věstonice. PLoS ONE 11, e0163019 (2016).
Lamnidis, T. C. et al. Ancient Fennoscandian genomes reveal origin and spread of Siberian ancestry in Europe. Nat. Commun. 9, 5018 (2018).
Korneliussen, T. S., Albrechtsen, A. & Nielsen, R. ANGSD: analysis of next generation sequencing data. BMC Bioinformatics 15, 356 (2014).
Renaud, G., Slon, V., Duggan, A. T. & Kelso, J. Schmutzi: estimation of contamination and endogenous mitochondrial consensus calling for ancient DNA. Genome Biol. 16, 224 (2015).
Fu, Q. et al. A revised timescale for human evolution based on ancient mitochondrial genomes. Curr. Biol. 23, 553–559 (2013).
Skoglund, P. et al. Separating endogenous ancient DNA from modern day contamination in a Siberian Neandertal. Proc. Natl Acad. Sci. USA 111, 2229–2234 (2014).
Lazaridis, I. et al. Genomic insights into the origin of farming in the ancient Near East. Nature 536, 419–424 (2016).
Lazaridis, I. et al. Ancient human genomes suggest three ancestral populations for present-day Europeans. Nature 513, 409–413 (2014).
Li, H. et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Behar, D. M. et al. A ‘Copernican’ reassessment of the human mitochondrial DNA tree from its root. Am. J. Hum. Genet. 90, 675–684 (2012).
Kearse, M. et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649 (2012).
Weissensteiner, H. et al. HaploGrep 2: mitochondrial haplogroup classification in the era of high-throughput sequencing. Nucleic Acids Res. 44, W58–W63 (2016).
Kennett, D. J. et al. Archaeogenomic evidence reveals prehistoric matrilineal dynasty. Nat. Commun. 8, 14115 (2017).
Monroy Kuhn, J. M., Jakobsson, M. & Günther, T. Estimating genetic kin relationships in prehistoric populations. PLoS ONE 13, e0195491 (2018).
Lipatov, M., Sanjeev, K., Patro, R. & Veeramah, K. R. Maximum likelihood estimation of biological relatedness from low coverage sequencing data. Preprint at bioRxiv https://doi.org/10.1101/023374 (2015).
Sudmant, P. H. et al. An integrated map of structural variation in 2,504 human genomes. Nature 526, 75–81 (2015).
Yunusbayev, B. et al. The Caucasus as an asymmetric semipermeable barrier to ancient human migrations. Mol. Biol. Evol. 29, 359–365 (2012).
1000 Genomes Project Consortium et al. A global reference for human genetic variation. Nature 526, 68–74 (2015).
Behar, D. M. et al. The genome-wide structure of the Jewish people. Nature 466, 238–242 (2010).
Kushniarevich, A. et al. Genetic heritage of the Balto-Slavic speaking populations: a synthesis of autosomal, mitochondrial and Y-chromosomal data. PLoS ONE 10, e0135820 (2015).
Pagani, L. et al. Genomic analyses inform on migration events during the peopling of Eurasia. Nature 538, 238–242 (2016).
Kovacevic, L. et al. Standing at the gateway to Europe-the genetic structure of western Balkan populations based on autosomal and haploid markers. PLoS ONE 9, e105090 (2014).
Price, A. L. et al. Long-range LD can confound genome scans in admixed populations. Am. J. Hum. Genet. 83, 132–135; author reply 135–139 (2008).
Anderson, C. A. et al. Data quality control in genetic case–control association studies. Nat. Protoc. 5, 1564–1573 (2010).
Purcell, S. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575 (2007).
Patterson, N., Price, A. L. & Reich, D. Population structure and eigenanalysis. PLoS Genet. 2, e190 (2006).
Patterson, N. et al. Ancient admixture in human history. Genetics 192, 1065–1093 (2012).
Pickrell, J. K. & Pritchard, J. K. Inference of population splits and mixtures from genome-wide allele frequency data. PLoS Genet. 8, e1002967 (2012).
Reich, D. et al. Reconstructing Native American population history. Nature 488, 370–374 (2012).
Acknowledgements
We thank the Brighton & Hove Museums and A. Maxted; the Stiftung Archäologie im rheinischen Braunkohlerevier; the Max Planck Society; N. Adamski and A. Claxton; and the Velux Foundations. This project has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement number 851511). P.J. and K.D. were supported by a Leverhulme Doctoral Scholarship awarded to M.B.R. and M.P. C.J.E. acknowledges support from The Leverhulme Trust via research project grant RPG-388. I.B., T. Booth, M.G.T. and S. Brace were supported by a Wellcome Trust Investigator Award (project number 100713/Z/12/Z). S. Beckett and M.V.T. thank R. Baldry and North West Norfolk History Society for funding of radiocarbon dating analyses. Sampling and DNA extraction of the samples from Groningen was funded by the European Funds for Regional Development and the Province of Groningen. We are grateful to the Museum für Archäologie in der Stiftung Schleswig-Holsteinische Landesmuseen Schloss Gottorf for providing samples from Schleswig Rathausmarkt. The ancient DNA laboratory work at Harvard University was supported by the US National Institutes of Health grant GM100233, by John Templeton Foundation grant 61220, by a gift from J.-F. Clin, by the Howard Hughes Medical Institute, and by the Allen Discovery Center program, a Paul G. Allen Frontiers Group advised program of the Paul G. Allen Family Foundation.
Funding
Open access funding provided by Max Planck Society.
Author information
Authors and Affiliations
Contributions
E.A., C.J.E., S.J., L. Lacher, S. Sabin, K. Schmidt, M.A.B., S.K.-B., F.B., D.W., D.N., L.S., S.K., S. Beckett, M.V.T., N.F., C.R., T. Barton, J.C., J.H., U.W., H.D., J.J.C., A.R., E.P., N.D., S.L., T.P., R.M., F.S., D. Swales, A.S., D.P., R.K., L. Ladle, C.P., S.G.-P., P.P., R.D., C.S., J.W., A.M.J., A.V.R., R.S. and A.C.R. provided samples and archaeological contextualization. J.G., P.J., E.A., K.D., C.J.E., S.J., L. Lacher, S. Sabin, W.H., R.R., S. Brace, M.A.B., N.B., K.C., E.H., L.I., A.M.L., M.M., K. Stewardson and F.Z. performed the laboratory work. J.G., D. Sayer, P.J., M.P., S.S.E., K.H.S.M., N.P., L.P., D.R. and S. Schiffels analysed the genetic data. M.P., C.J.E., Å.J.V., N.R., B.K.-K., V.J.S., I.B., A.C., K.I.B., T. Booth, H.S., M.G.T., A.H., M.B.R., D.R., J.K. and S. Schiffels supervised the laboratory work, sampling or analysis. J.G., D. Sayer and S. Schiffels wrote the paper with input from all coauthors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Daniel Bradley, Helena Hamerow and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Genetic affinity statistics between ancient and present-day northwestern European populations.
a) FST between relevant present-day Europeans and England_EMA (n = 285). Populations are coloured to their respective language family affiliation. Belgium is classified as Germanic, since Belgian samples were recruited mainly amongst patients from the northern Flemish-speaking region of Belgium. Error bars represent ± 3 standard errors. Samples sizes for present-day European populations are indicated in Supplementary Table 3.1 b) FST between relevant present-day Europeans and England_IA (n = 290). Error bars represent ± 3 standard errors. c) F-statistics of the form F4(YRI, TestA; Ireland, TestB). Negative values indicate that the test population is closer to Ireland than to TestB; positive values indicate that the test population is closer to TestB than to Ireland. d) Same for the F-statistics of the form F4(YRI, TestA; Denmark, TestB).
Extended Data Fig. 2 Individual-based ancestry decomposition and population affinities through time.
a) f-statistics of the form F4(YRI, Test; WBI, CNE) for 758 ancient English individuals. Data are presented as point estimates for the respective F4-statistic ± 2 standard errors. Negative values indicate that the test individual is closer to WBI than to CNE; positive values indicate that the test population is closer to CNE than to WBI. b) Modelling ancient post-Neolithic individuals from England and Ireland as a mixture of CNE individuals (red) and the WBI individuals (blue). Data are presented as admixture proportions. Each bar represents genome-wide mixture proportions for one individual. Individuals are ordered chronologically.
Extended Data Fig. 3 Regional ancestry decomposition and population affinities through time.
a) for England: left) f-statistics of the form F4(YRI, Test; WBI, CNE). Data are presented as exact F4-values. Negative values indicate that the test population is closer to WBI than to CNE; positive values indicate that the test population is closer to CNE than to WBI. Error bars represent ± 2 standard errors. Samples sizes for Test populations are indicated in Supplementary Table 3.3 right) Mean CNE and WBI ancestry proportions per period as inferred using supervised ADMIXTURE. Error bars represent ± 1 standard error of the mean. b) same for Scotland. c) same for Wales. d) same for Ireland.
Extended Data Fig. 4 Family tree reconstruction featuring integration of local ancestry into an immigrant kin group.
a) The genetic pedigree of 13 related individuals at Dover Buckland. Indicated are the mtDNA haplogroups, Y-chromosome haplogroups, and associated grave goods of each individual. Males are depicted as squares, females as circles. b) Spatial distribution of the addressed burials across the site. Genetically related burials are connected with lines. c) Genetic distribution of the addressed individuals across a Principal Components Analysis of present-day genomes from northwestern Europe. Genetically related individuals are connected with lines.
Extended Data Fig. 5 Visualisations of genetic affinity between early medieval English individuals and contemporary continental populations.
a) Multidimensional Scaling Plot of pairwise F3 distances of the form F3(CHB, ancient population A, ancient population B). b) PCA of F4 statistics of the form F4(YRI, ancient population; TestA, TestB). TestA and TestB iterate through 15 present-day European populations (Methods). Additionally, a neighbour-joining tree of the same dataset was projected onto the two PCs. England_EMA_CNE are those early medieval English individuals who have exclusively CNE ancestry.
Extended Data Fig. 6 Measures of CNE ancestry in early medieval English sites.
a) F4 statistics of the form F4(YRI, Site; WBI, CNE). Data are presented as point estimates ± 2 standard errors. Samples sizes for early medieval English sites are indicated in Supplementary Table 5.3 b) Mean supervised ADMIXTURE proportions at K = 2. Error bars represent ± 1 standard error of the mean. c) qpAdm admixture proportions and p-values using a two-way admixture model of England_EMA with England_LIA_Roman (n = 32) and LowerSaxony_EMA (n = 39) as sources. Error bars represent ± 1 standard error.
Extended Data Fig. 7 Extended Principal Components Analysis.
a) Principal Components Analysis of present-day genomes from northwestern Europe. IE = Northern Ireland & Ireland, WA = Wales, SC = Scotland, ENG = England, NL = Netherlands, GER = Northern Germany, DK = Denmark, NO = Norway, SE = Sweden, BE = Belgium, FR = France. b) Genetic structure of published and novel ancient individuals in this study, projected onto a). Polygons indicate where 2/3 of the data is located (England BA+IA, North Sea IA+EMA, and France IA+WGermany EMA, respectively). The Scandinavian Iron Age samples are connected with lines for clarity. Western Germany comprises samples from Alt-Inden; Northern Germany comprises samples from Lower Saxony, Mecklenburg-Vorpommern, and Schleswig-Holstein; Denmark_MA comprises samples from Copenhagen. BA = Bronze Age (EBA, MBA, and LBA = early, middle, and late BA), IA = Iron Age, RA = Roman Age, MA = Middle Ages (EMA=early MA), For rough time boundaries of the samples used here, see Methods.
Extended Data Fig. 8 Correlations of early medieval ancestry with archaeological and linguistic evidence.
a) Distribution of early Anglo-Saxon cemeteries, Celtic river names, and Frankish objects. Ancestry proportions from supervised ADMIXTURE at K = 3 are shown for early Anglo-Saxon sites. CNE ancestry is shown in red, WBI in blue, and CWE in green. b) qpAdm ancestry proportions for 31 present-day populations from Britain and Ireland using LowerSaxony_EMA as a source for CNE ancestry. Number of individuals used for each source is indicated (England_LIA, LowerSaxony_EMA, and France_IA; n = 32, 39, and 26, respectively). Samples sizes for present-day PoBI sampling regions are indicated in Supplementary Table 5.16. Error bars represent ± 1 standard error.
Supplementary information
Supplementary Information
This file contains Supplementary Notes 1-7. The file also includes Supplementary Figs. 1.1–7.1, Supplementary Tables 2.2–2.7 and additional references.
Supplementary Data 1
This Excel file contains the Supplementary Tables 1, 2.1, and 2.8–7.1.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gretzinger, J., Sayer, D., Justeau, P. et al. The Anglo-Saxon migration and the formation of the early English gene pool. Nature 610, 112–119 (2022). https://doi.org/10.1038/s41586-022-05247-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05247-2
This article is cited by
-
Windows into the past: recent scientific techniques in dental analysis
British Dental Journal (2024)
-
The Allen Ancient DNA Resource (AADR) a curated compendium of ancient human genomes
Scientific Data (2024)
-
Genetic history of East-Central Europe in the first millennium CE
Genome Biology (2023)
-
Unappreciated subcontinental admixture in Europeans and European Americans and implications for genetic epidemiology studies
Nature Communications (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
