
Abstract
The neural cell adhesion molecule (NCAM) is a glycoprotein implicated in cell–cell adhesion, neurite outgrowth and synaptic plasticity. Polysialic acid (polySia) is mainly attached to NCAM (polySia-NCAM) and has an essential role in regulating NCAM-dependent developmental processes that require plasticity, that is, cell migration, axon guidance and synapse formation. Post-mortem and genetic evidence suggests that dysregulation of polySia-NCAM is involved in schizophrenia (SZ). We enrolled 45 patients diagnosed with SZ and 45 healthy individuals who were submitted to polySia-NCAM peripheral quantification, cognitive and psychopathological assessment and structural neuroimaging (brain volumes and diffusion tensor imaging). PolySia-NCAM serum levels were increased in SZ patients, independently of antipsychotic treatment, and were associated with negative symptoms, blunted affect and declarative memory impairment. The increased polySia-NCAM levels were associated with decreased volume in the left prefrontal cortex, namely Brodmann area 46, in patients and increased volume in the same brain area of healthy individuals. As this brain region is involved in the pathophysiology of SZ and its associated phenomenology, the data indicate that polySia-NCAM deserves further scrutiny because of its possible role in early neurodevelopmental mechanisms of the disorder.
Similar content being viewed by others
Introduction
Development of brain connectivity requires temporal and spatial control of cellular interactions. The neural cell adhesion molecule (NCAM) is a member of the immunoglobulin superfamily and is a key factor in these processes, including axonal/dendritic growth and branching and synaptic plasticity.1 In vertebrates, NCAM can be modified by polysialic acid (polySia), a linear homopolymer of sialic acids that is mainly present during fetal and early postnatal brain development.2, 3 PolySia disappears almost completely from the adult brain, except for some areas of continuous neurogenesis and plasticity, whereas NCAM expression levels remain relatively unchanged. When polySia is present on NCAM it acts as a negative regulator of cell interactions associated with a variety of developmental processes that require plasticity, including cell migration, the guidance and targeting of axons and synapse formation.2, 3 The loss of polySia-NCAM (polySia attached to NCAM) in NCAM-deficient mice or in wild-type mice treated with a polySia-specific endosialidase has been associated with alterations in a variety of brain functions, including learning and memory behaviors and both long-term potentiation and long-term depression in the hippocampus.4 Imbalanced synthesis of polySia and NCAM causes pathological brain development, including reduction of prefrontal cortex (PFC) interneurons, hypoplasia of major brain axon tracts, enlarged ventricles, reduced size of the thalamus and disturbed thalamus–cortex connectivity, which may be linked to the deficits in the cognitive performance observed in these mice.5, 6, 7 Furthermore, a synergistic negative consequence of polySia deficiency and cannabis exposure on cognitive performance has been demonstrated,8 and in a double-hit animal model of schizophrenia (SZ) prefrontal changes of polySia were detected together with altered excitatory–inhibitory balance.9
SZ is a mental disorder that is intimately related to neurodevelopmental and neurodegenerative conditions involving disturbed long-range and synaptic connectivity.10, 11 Notably, these abnormalities occur during a very restricted stage of brain development. A number of studies indicate a link between dysregulation of the polySia-NCAM system and SZ.1, 9, 12, 13 Altered polySia-NCAM immunoreactivity was observed in schizophrenic hippocampi14 and dorsolateral PFC.9 This is particularly intriguing because impairment of hippocampal and PFC functions as well as disturbances in their anatomical organization are also involved in the etiology of SZ.15, 16 Altered concentrations of NCAM isoforms and soluble NCAM fragments have been frequently observed in the cerebrospinal fluid of SZ patients,1 as well as in the hippocampus, PFC and other cortical areas including Brodmann area 46.17 Genetic association studies have also identified NCAM and the polySia-synthesizing enzyme ST8SIA2 as a candidate susceptibility gene for SZ; however, findings are not fully consistent.9, 12, 13, 18, 19 Taken together, these findings raise the question of whether polySia-NCAM has a causal role in the development of SZ symptoms.
Because dysregulation of polySia-NCAM accounts for a long-range hypoconnectivity, reduced gray matter (GM) volumes and cognitive deficits in the mouse model,5, 7 we hypothesized that polySia-NCAM levels in SZ might be linked to brain structural variations and clinical parameters of cognition and psychopathology. To test this hypothesis, serum levels of polySia-NCAM were assessed in a group of 45 SZ patients and 45 age- and gender-matched healthy controls (HC), and were correlated to macrostructural (volume) and microstructural (diffusion tensor-derived) measures of brain structural integrity as well as to psychopathological and cognitive symptoms in the patient group.
Materials and methods
Subjects
As no preliminary data comparing polySia-NCAM between HC and SZ are available, sample numerosity was chosen assuming a Type I error rate α=0.05, a medium-high Cohen’s effect size of 0.6 and a statistical power of 0.8.20 With these assumptions the size of the total sample, HC plus SZ, results to be 90.
For this study we initially included 62 SZ patients consecutively recruited at IRCCS Santa Lucia Foundation of Rome. The diagnosis of SZ was made according to the Diagnostic and Statistical Manual of Mental Disorders IV-Edition, text revised (DSM-IV-TR).21 The clinician who had been treating the patients and knew their clinical history, but who was blind to the aims of the study, made the preliminary diagnosis. Then, a senior research psychiatrist confirmed all preliminary diagnoses using the Structured Clinical Interview for DSM-IV-TR-Patient Edition (SCID-I/P).22 Out of the original group of patients confidently diagnosed with SZ, four refused to undergo the magnetic resonance imaging exam, seven were excluded for strong movement artifacts in brain images and six were excluded because of the presence of moderate to severe brain vascular lesions (see exclusion criteria below). The remaining 45 SZ patients were age- and gender-matched with 45 HC consecutively recruited from universities, community recreational centers and hospital personnel through local advertisement.
Overall severity of psychiatric symptoms was assessed using the Positive and Negative Syndrome Scale (PANSS).23 Age at onset was defined as the age at onset of positive or negative symptoms preceding the first hospitalization, which was investigated in an interview with patients and first-degree relatives.
All patients, but six, were receiving stable oral dosages of one or more atypical antipsychotics such as risperidone, quetiapine and olanzapine. Antipsychotic dosages were converted to equivalents of chlorpromazine.24 Extrapyramidal side effects and abnormal involuntary movements were evaluated using the Simpson Angus Scale25 and the Abnormal Involuntary Movement Scale.26
All HCs were screened for a current or lifetime history of DSM-IV-TR Axis I and II disorders using the SCID-I/NP and SCID-II;27 they were also assessed to confirm that no first-degree relative had a history of psychosis.
Inclusion criteria for all participants were as follows: (1) age between 18 and 65 years, (2) at least 8 years of education and (3) suitability for magnetic resonance imaging scanning. Exclusion criteria were as follows: (1) history of alcohol or drug abuse in the 2 years before the assessment, (2) lifetime drug dependence, (3) traumatic head injury with loss of consciousness, (4) past or present major medical illness or neurological disorders, (5) any (for HC) or additional (for SZ) psychiatric disorder or mental retardation, (6) dementia or cognitive deterioration according to DSM-IV-TR criteria and a Mini-Mental State Examination (MMSE)28 score lower than 25, consistent with normative data in the Italian population29 and (7) any potential brain abnormality and microvascular lesion as apparent on conventional FLAIR scans; in particular, the presence, severity and location of vascular lesions were computed according to the semiautomated method recently published by our group.30
Sociodemographic and clinical characteristics of the SZ and HC samples are shown in Table 1.
The study was approved and undertaken in accordance with the guidelines of the Santa Lucia Foundation Ethics Committee. All participants gave their written informed consent to participate after they had received a complete explanation of the study procedures.
Determination of polySia-NCAM serum levels
Sera were obtained by centrifugation of clotted blood samples, and aliquots were stored at −80 °C. For determination of polySia-NCAM, serum samples were thawed and assayed using a sandwich enzyme-linked immunosorbent assay (modified from Takamatsu et al.31 and detailed in Supplementary Materials). Each sample was analyzed by at least four independent experiments and values were normalized to the mean value of all measured samples (100%).
Psychopathological and cognitive assessment
The PANSS23 was administered to rate the severity of psychopathological symptoms. The PANSS rates the patient from 1 to 7 on 30 different symptoms based on the interview as well as reports of family members or primary care hospital workers. The symptoms are grouped into three global scales, that is, Positive Symptoms Scale, Negative Symptoms Scale and General Psychopathology Scale, and five subscales, that is, Paranoid-Belligerence, Anergia, Depression, Activation and Thought Disturbance. As 1, rather than 0, is given as the lowest score for each item, a patient cannot score lower than 30 for the total PANSS score. PANSS ratings were obtained on all information available pertaining to the last week of the assessment. Aggressive behavior was assessed through the administration of the Modified Overt Aggression Scale.32
With regard to the neuropsychological assessment, the MMSE was administered to obtain a global index of cognitive deterioration. Several tests were selected to provide information about the functioning of different cognitive domains such as: verbal memory (Mental Deterioration Battery (MDB) Rey’s 15-word Immediate Recall and Delayed Recall); short-term visual memory (MDB Immediate Visual Memory); logical reasoning (MDB Raven’s Progressive Matrices’ 47); simple constructional praxis (MDB Copying Drawings and Copying Drawings with Landmarks); language (MDB Phonological Verbal Fluency and Category Fluency);33 executive functions (Modified Wisconsin Card Sorting test);33 and divided attention and attentional control (Double Barrage test).34 We also administered the Rey–Osterrieth Complex Figure Test immediate copy35 to evaluate visuo-constructive abilities.
Image acquisition and processing
All the 90 participants underwent the same imaging protocol, which included three-dimensional T1-weighted, DTI, T2-weighted and FLAIR sequences, using a 3T Allegra MR imager (Siemens, Erlangen, Germany) with a standard quadrature head coil.
T1 and DTI images were processed in order to obtain GM and white matter (WM) volumetric maps as well as fractional anisotropy and mean diffusivity maps. Details about the image-processing methodology can be found in the Supplementary Materials.
Statistical analyses—clinical and polySia-NCAM
Differences between diagnostic groups on sociodemographic, clinical and biological variables were assessed by means of unpaired t-tests. Relationships among continuous variables were analyzed by Pearson’s product moment correlations (Fisher’s r to z transformations). The statistical threshold was set at P<0.05.
Statistical analyses—neuroimaging
In order to avoid possible edge effects between different tissue types, the voxel-based morphometry analyses of GM and WM volumes were carried out by excluding all voxels that had a less than 20% probability of belonging to the relative tissue (absolute threshold masking). Further, analyses of mean diffusivity maps were restricted to cortical and deep GM structures by means of an inclusive mask obtained by averaging subjects’ GM segments and excluding all voxels with a less than 30% probability of belonging to GM. Finally, statistical analyses on fractional anisotropy maps were restricted to voxels in the WM skeleton.
Although a parametric approach within the SPM8 framework was used for volumetric data, DTI analyses were carried out with a nonparametric permutation-based methodology (that is, the randomize tool, implemented in FSL).
The differential relationship between polySia-NCAM and neuroimaging parameters in the two HC and SZ groups was assessed as follows: subjects’ polySia-NCAM levels were entered in four analysis of covariance designs (one for each parameter) as continuous variables, thus modeling the main effect of group (treated as independent variable) and its interaction with polySia-NCAM levels on each neuroimaging parameter (GM and WM volume, GM mean diffusivity and WM tracts fractional anisotropy). Positive (polySia-NCAMHC–polySia-NCAMSZ) and negative (polySia-NCAMSZ–polySia-NCAMHC) interaction contrasts were defined to assess the differential relationship between polySia-NCAM levels and neuroimaging parameters in the two HC and SZ groups. MMSE scores were entered in the model as nuisance variable.
The statistical threshold was set at P<0.05 using a family-wise error correction that accounted for multiple comparisons and a cluster-size threshold of 30 contiguous voxels. The mean volumetric values of the brain areas, where significant effects were found, were extracted for each subject using an in-house shell script. These values were subsequently used to create scatterplots showing the direction of the relationship.
Results
As shown in Table 1, the two groups were perfectly matched for gender distribution (31 male subjects in both groups) and did not differ for age; HC had more years of formal education and higher MMSE scores.
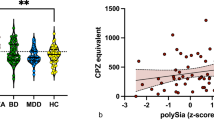
SZ showed higher levels of polySia-NCAM. This did not seem to be driven by treatment because no correlation was found between polySia-NCAM levels and antipsychotic drug dosages in equivalents of chlorpromazine (r=0.024; P=0.822). We also computed the relationship between polySia-NCAM levels and severity of positive and negative symptoms as well as cognitive performance of the SZ sample. We found a significant positive correlation among polySia-NCAM levels, the PANSS-negative scale score (r=0.317; P=0.033) and the PANSS-blunted affect score (r=0.304; P=0.041) as well as a significant negative correlation between polySia-NCAM and delayed recall of the Rey–Osterrieth Complex Figure Test score (r=−0.321; P=0.032). No significant correlations were found between polySia-NCAM levels and positive symptoms as well as Modified Overt Aggression Scale scores.
With respect to the neuroimaging analyses, we found a statistically significant (P<0.05, family-wise error-corrected) interaction between the diagnostic group variable and polySia-NCAM on GM volume in a prefrontal cortical area. Namely, in a cluster located in the left PFC (Montreal Neurological Institute coordinates: x=−16; y=36; z=27; Brodmann area 46; Z-score=4.68; extent: 218 voxels), where SZ patients showed a negative relationship between GM volume and polySia-NCAM levels, whereas HC showed a positive association (see Figure 1a).

Relationship between neural cell adhesion molecule (NCAM) serum levels and gray matter volume in healthy control (HC) and schizophrenia (SZ). The statistical result (red-to-yellow map) is superimposed over a three-dimensional rendering of the Montreal Neurological Institute standard brain. A scatterplot shows the differential relationship between NCAM and volume in the two study groups.
Regarding WM volumetric analyses, as well as WM fractional anisotropy and GM mean diffusivity, no results survived the P<0.05, family-wise error-corrected, threshold.
Discussion
The aim of the present study was to investigate whether polySia-NCAM serum levels in SZ patients are different from those of HC and whether they are associated with brain macro- and microstructural variations and psychopathological-cognitive symptomatology. To the best of our knowledge, this is the first study investigating this issue and, therefore, our work could provide an additional piece for the complex puzzle of research on the biological mechanisms implied in SZ pathogenesis. SZ is a psychiatric disorder with a complex pathophysiology that is influenced by multiple factors. It is believed that SZ is highly related to both neurodevelopmental and neurodegenerative processes involving disconnectivity and disorder of synapses.36 Thus, several factors are intricately involved in the pathophysiology of SZ and make it difficult to decipher the underlying mechanisms. Nevertheless, the unprecedented correlations of polySia-NCAM serum levels with brain structural variation and, at the same time, with negative symptoms of schizophrenic patients (as demonstrated by the current study) strongly suggest that dysregulation of polySia-NCAM is an important molecular mechanism in the pathophysiology of the disorder.
We found that polySia-NCAM levels were more elevated in SZ patients when compared with HC and that they were differently associated with GM volume in the two groups. Namely, increased polySia-NCAM levels were linked to greater volumes in the HC sample and were associated with reduced volumes in SZ patients. This reverse relationship of polySia-NCAM serum levels and brain integrity was found in a cortical area (that is, the left PFC (Brodmann area 46)), whose early alteration is one of the most replicated findings in SZ. In particular, as recently reviewed by Thermenos et al.,37 there is significant evidence of reduced PFC volume in high-risk and early SZ patients that becomes more significant over time and is associated with higher symptom severity. Post-mortem studies are consistent with the likelihood of primary structural abnormalities, possibly accounting for alteration of the PFC in SZ. Indeed, findings of increased cell packing density without a change in neuronal number suggest that PFC volume reduction may be linked to a decreased amount of neuropil in SZ.38, 39, 40, 41 This is mirrored by functional alteration in the PFC observed across a variety of cognitive tasks (for example, working memory, semantic encoding and language tasks), symptoms and variance in SZ risk genes. Such PFC abnormalities seem to be associated with developmental processes that could be linked to polySia-NCAM alterations. In fact, it has been suggested that abnormal early neurodevelopment (whether linked to genetic or environmental factors or both) may impair the process of normal maturational changes that occurs in adolescence (for example, synaptic remodeling,42 WM development43 and sensitivity to effects of stress and sex hormones44). The final development of the PFC in late adolescence and young adulthood45 may be particularly at risk, with adverse implications for both PFC structure and function including maturation of activity, connectivity and network topology.46, 47 To date, a range of evidence suggests that macroscopic changes in GM volume over time are not because of the degeneration of neurons, but may reflect (at least in part) the end point of a longstanding disturbance in the plastic rearrangement of the neural connectivity architecture, especially at the level of synapses and prefrontal cortical connections.48 In this view, the altered expression of polySia-NCAM may be linked to neurodevelopment and neuroplasticity processes, ultimately leading to changes in the volume of brain areas such as the PFC, where altered concentrations of polySia and NCAM isoforms have been previously found in SZ patients.9, 17 In summary, because neurodevelopmental mechanisms affect brain structure before the onset of psychosis and continue throughout the progression of the disease, the polySia-NCAM changes may not be involved only in neurodevelopmental mechanisms, as evident from animal models.49 Ongoing dysregulation of polySia-NCAM, as evidenced by the correlation of polySia-NCAM serum levels with altered brain structure in SZ but not HC, may also be relevant for the mechanisms that cause the disease-specific progression of brain structural changes.
Further, coherently with imaging data we found that polySia-NCAM levels were associated with increased negative symptoms and blunted affect (that is, diminished emotional responsiveness, as characterized by a reduction in facial expression, modulation of feelings and communicative gestures) as well as impaired long-term visual–spatial memory. These findings are in line with several previous reports in both animal and human studies. As stated, polySia-NCAM is involved in several neurodevelopmental processes and has been suggested to have a role in the negative and cognitive symptoms of SZ, which are known to present early in the course of the illness.49 For instance, NCAM−/− mice display a 10% reduction in the overall brain weight and a 36% decline in the size of the olfactory bulb as well as impaired spatial learning.50 Using NCAM-180 knockout mice, Wood et al.51 demonstrated a marked increase in both the left and right anterior horns of the lateral ventricles and reduced prepulse inhibition, a behavioral model of SZ. Furthermore, reduced polysialylation causes pathological brain development comprising reductions of parvalbumin-positive interneurons in the PFC6 as well as enlarged lateral ventricles, smaller thalamus and reduced thalamocortical connectivity together with impaired working memory and prepulse inhibition.7
Conversely to negative symptoms, in our sample polySia-NCAM serum levels did not result to be linked to positive symptomatology or aggressive behavior, whereas previous animal studies using both deficient and postnatally treated mice showed a link between polySia-NACM defect and increased aggressive behavior.52, 53 A possible rationale accounting for this discrepancy relies in the fact that in these previous studies stress was usually experimentally manipulated, and resulted to significantly interact with polySia-NCAM decreased expression in modulating aggressive and emotional behavior.
The increased polySia-NCAM serum levels in SZ patients, such as those detected in the current study, contrast with reports of lower numbers of polySia-positive cells in the hilus region of the hippocampus and reduced polySia immunoreactivity in the PFC of SZ patients.9, 14 This discrepancy in results may be explained by previous results. For NCAM, an inverse relationship between levels found in the brain and in the serum was reported in Alzheimer’s disease. In particular, serum NCAM levels were increased in patients with high Global Deterioration Scale values,54 whereas they were decreased in the frontal and temporal cortex55 and significantly fewer NCAM-positive neurons were detected in the frontal cortex.56 Therefore, it may well be that the higher polySia-NCAM serum levels in SZ are linked to altered expression in the brain, possibly reflecting neurodegenerative processes.
Similar to the results of the current study, Lyons et al.57 found an increase in the NCAM serum levels in SZ patients when compared with normal controls and demonstrated that this difference was more marked in type II SZ (that is, in those patients who showed more negative symptoms). In contrast, a recent study detected significantly lower NCAM serum levels in SZ patients.58 Differences in the efficiency of capturing and/or detecting different NCAM species could be responsible for these seemingly divergent results. Furthermore, it is well known that recognition of NCAM is strongly reduced in the presence of polySia.59 Hence, the elevated polySia-NCAM levels of SZ patients found in the current study could cause reduced NCAM detection, as observed by Zhang et al.58 In addition to these reciprocal findings for NCAM and polySia-NCAM, previous report indicated that the serum levels are inversely related to those found in the brains of SZ patients. Also, reduced polySia immunoreactivity in the PFC and hippocampus, as discussed above, contrasts with an increase in NCAM fragments in the same brain areas.60 Furthermore, in a study on a large sample of SZ patients (n=641) Sullivan et al.61 found an association between five contiguous single-nucleotide polymorphisms (rs1943620, rs1836796, rs1821693, rs686050 and rs584427, associated with gene NCAM1) and cognitive functioning. Thus, the present data seem to support the view that polySia-NCAM abnormalities in SZ are strongly linked to volume reduction in PFC and, coherently, with negative symptoms and cognitive disorders.
SZ is a complex disease in which both biological and environmental factors contribute to the pathogenesis. It is therefore likely that the mechanisms through which altered polySia-NCAM serum levels are connected to the disorder are also complex. Nevertheless, altered serum levels, such as those detected in the current study, may be indicative of disturbed polySia-NCAM homeostasis, which, as discussed above, has the potential to profoundly affect brain development and connectivity. The clear correlations of polySia-NCAM serum levels with altered brain structural measures underscore this link. In parallel, the intriguing result of a positive correlation between polySia-NCAM and prefrontal volume in the HC sample claims in favor of the proven relationship between polySia-NCAM expression and adult neuroplasticity. This is suggested by the remarkable amounts within the main adult neurogenic areas such as the subventricular zone, the olfactory bulb and the dentate gyrus, as well as by the reciprocal relationship between polySia-NCAM and learning and memory.62
A final point merits further discussion, namely the absence of correlation between polySia-NCAM levels and antipsychotic drug dosages. This is in partial disagreement with previous findings showing a significant and selective relationship between polySia-NCAM and antipsychotic treatment. In particular, Frasca et al.63 performed a 2-week treatment with the first-generation antipsychotic haloperidol and the second-generation antipsychotic olanzapine and analyzed the expression of polySia-NCAM in the rat hippocampus and PFC via immunohistochemistry. Results showed a regional- and drug-selective increase in the polySia-NCAM expression in PFC of olanzapine-treated rats with no effects in the hippocampus; conversely, haloperidol did not produce a change in either brain region. However, these results are hardly comparable to those obtained in our study for several reasons. First, rats used in the work of Frasca et al. were not animal model of SZ and, as such, results on polySia-NCAM expression can only be linked to the influence of treatment and not to the possible role of the disease per se. Second, our SZ sample was composed of patients with long duration of illness and thus were chronically treated, whereas rats received a 2-week treatment. Therefore, it could be the case that the results of Frasca et al. highlight the early influence of antipsychotic treatment on polySia-NCAM expression thus overlooking potential long-term effects. Future studies should be specifically designed to unravel this key issue.
In conclusion, as demonstrated by the present study, exploring the mechanisms that establish the link between altered polySia-NCAM levels and behavioral and neuroimaging data should provide further insight into SZ diagnosis, prognosis and treatment.
References
Brennaman LH, Maness PF . NCAM in neuropsychiatric and neurodegenerative disorders. Adv Exp Med Biol 2010; 663: 299–317.
Rutishauser U . Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat Rev Neurosci 2008; 9: 26–35.
Schnaar RL, Gerardy-Schahn R, Hildebrandt H . Sialic acids in the brain: gangliosides and polysialic acid in nervous system development, stability, disease, and regeneration. Physiol Rev 2014; 94: 461–518.
Becker CG, Artola A, Gerardy-Schahn R, Becker T, Welzl H, Schachner M . The polysialic acid modification of the neural cell adhesion molecule is involved in spatial learning and hippocampal long-term potentiation. J Neurosci Res 1996; 45: 143–152.
Hildebrandt H, Mühlenhoff M, Oltmann-Norden I, Röckle I, Burkhardt H, Weinhold B et al. Imbalance of neural cell adhesion molecule and polysialyltransferase alleles causes defective brain connectivity. Brain 2009; 132: 2831–2838.
Kröcher T, Röckle I, Diederichs U, Weinhold B, Burkhardt H, Yanagawa Y et al. A crucial role for polysialic acid in developmental interneuron migration and the establishment of interneuron densities in the mouse prefrontal cortex. Development 2014; 141: 3022–3032.
Kröcher T, Malinovskaja K, Jürgenson M, Aonurm-Helm A, Zharkovskaya T, Kalda A et al. Schizophrenia-like phenotype of polysialyltransferase ST8SIA2-deficient mice. Brain Struct Funct 2015; 220: 71–83.
Tantra M, Kröcher T, Papiol S, Winkler D, Röckle I, Jatho J et al. St8sia2 deficiency plus juvenile cannabis exposure in mice synergistically affect higher cognition in adulthood. Behav Brain Res 2014; 275: 166–175.
Gilabert-Juan J, Varea E, Guirado R, Blasco-Ibáñez JM, Crespo C, Nácher J . Alterations in the expression of PSA-NCAM and synaptic proteins in the dorsolateral prefrontal cortex of psychiatric disorder patients. Neurosci Lett 2012; 530: 97–102.
Schmitt A, Hasan A, Gruber O, Falkai P . Schizophrenia as a disorder of disconnectivity. Eur Arch Psychiatry Clin Neurosci 2011; 261: S150–S154.
Rapoport JL, Giedd JN, Gogtay N . Neurodevelopmental model of schizophrenia: update 2012. Mol Psychiatry 2012; 17: 1228–1238.
Arai M, Yamada K, Toyota T, Obata N, Haga S, Yoshida Y et al. Association between polymorphisms in the promoter region of the sialyltransferase 8B (SIAT8B) gene and schizophrenia. Biol Psychiatry 2006; 59: 652–659.
McAuley EZ, Scimone A, Tiwari Y, Agahi G, Mowry BJ, Holliday EG et al. Identification of sialyltransferase 8B as a generalized susceptibility gene for psychotic and mood disorders on chromosome 15q25-26. PLoS One 2012; 7: e38172.
Barbeau D, Liang JJ, Robitalille Y, Quirion R, Srivastava LK . Decreased expression of the embryonic form of the neural cell adhesion molecule in schizophrenic brains. Proc Natl Acad Sci USA 1995; 92: 2785–2789.
Adriano F, Caltagirone C, Spalletta G . Hippocampal volume reduction in first-episode and chronic schizophrenia: a review and meta-analysis. Neurosci 2012; 18: 180–200.
Zhou Y, Fan L, Qiu C, Jiang T . Prefrontal cortex and the dysconnectivity hypothesis of schizophrenia. Neurosci Bull 2015; 31: 207–219.
Gibbons AS, Thomas EA, Dean B . Regional and duration of illness differences in the alteration of NCAM-180 mRNA expression within the cortex of subjects with schizophrenia. Schizophr Res 2009; 112: 65–71.
Atz ME, Rollins B, Vawter MP . NCAM1 association study of bipolar disorder and schizophrenia: polymorphisms and alternatively spliced isoforms lead to similarities and differences. Psychiatr Genet 2007; 17: 55–67.
Tao R, Li C, Zheng Y, Qin W, Zhang J, Li X et al. Positive association between SIAT8B and schizophrenia in the Chinese Han population. Schizophr Res 2007; 90: 108–114.
Cohen J . Statistical Power Analysis for the Behavioral Sciences. Lawrence Erlbaum Associates Inc.: Hillsdale, NJ, England, 1977.
APA Diagnostic and Statistical Manual of Mental Disorders, 4th edn, American Psychiatric Association (APA): Washington, DC, USA, 2000.
First MB, Spitzer RL, Gibbon M, Williams JB . Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version, Patient Edition. (SCID-I/P). Biometrics Research, New York State Psychiatric Institute: New York, NY, USA, 2002.
Kay SR, Fiszbein A, Opler LA . The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull 1987; 13: 261–276.
Oquendo MA, Baca-Garcia E, Kartachov A, Khait V, Campbell CE, Richards M et al. A computer algorithm for calculating the adequacy of antidepressant treatment in unipolar and bipolar depression. J Clin Psychiatry 2003; 64: 825–833.
Simpson GM, Angus JW . A rating scale for extrapyramidal side effects. Acta Psychiatr Scand Suppl 1970; 212: 11–19.
Guy W. ECDEU Assessment Manual for Psychopharmacology, Revised. U.S. Dept. of Health, Education, and Welfare, Public Health Service, Alcohol, Drug Abuse, and Mental Health Administration, National Institute of Mental Health, Psychopharmacology Research Branch, Division of Extramural Research Programs: Rockville, MD, USA, 1976.
First MB, Gibbon M, Spitzer RL, Williams JB, Benjamin L . Structured Clinical Interview for DSM-IV Axis II Personality Disorders, (SCID-II). American Psychiatric Press Inc.: Washington, DC, USA, 1997.
Folstein MF, Folstein SE, McHugh PR . ‘Mini mental state’. A practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res 1975; 12: 189–198.
Measso G, Cavarzeran F, Zappalà G, Lebowitz BD, Crook TH, Pirozzolo FJ et al. The mini‐mental state examination: normative study of an Italian random sample. Dev Neuropsychol 1993; 9: 77–85.
Iorio M, Spalletta G, Chiapponi C, Luccichenti G, Cacciari C, Orfei MD et al. White matter hyperintensities segmentation: a new semi-automated method. Front Aging Neurosci 2013; 5: 76.
Takamatsu K, Auerbach B, Gerardy-Schahn R, Eckhardt M, Jaques G, Madry N . Characterization of tumor-associated neural cell adhesion molecule in human serum. Cancer Res 1994; 54: 2598–2603.
Kay SR, Wolkenfeld F, Murrill LM . Profiles of aggression among psychiatric patients. I. Nature and prevalence. J Nerv Ment Dis 1988; 176: 539–546.
Nyhus E, Barceló F . The Wisconsin Card Sorting Test and the cognitive assessment of prefrontal executive functions: a critical update. Brain Cogn 2009; 71: 437–451.
Carlesimo GA, Caltagirone C, Gainotti G . The Mental Deterioration Battery: normative data, diagnostic reliability and qualitative analyses of cognitive impairment. The Group for the Standardization of the Mental Deterioration Battery. Eur Neurol 1996; 36: 378–384.
Osterrieth PA . Le test de copie d’une figure complexe; contribution à l'étude de la perception et de la mémoire. / Test of copying a complex figure; contribution to the study of perception and memory. Arch Psychol (Geneve) 1944; 30: 206–355.
Chiapponi C, Piras F, Fagioli S, Piras F, Caltagirone C, Spalletta G . Age-related brain trajectories in schizophrenia: a systematic review of structural MRI studies. Psychiatry Res 2013; 14: 83–89.
Thermenos HW, Keshavan MS, Juelich RJ, Molokotos E, Whitfield-Gabrieli S, Brent BK et al. A review of neuroimaging studies of young relatives of individuals with schizophrenia: a developmental perspective from schizotaxia to schizophrenia. Am J Med Genet 2013; 162: 604–635.
Akbarian S, Bunney WE, Potkin SG, Wigal SB, Hagman JO, Sandman CA et al. Altered distribution of nicotinamide-adenine dinucleotide phosphate-diaphorase cells in frontal lobe of schizophrenics implies disturbances of cortical development. Arch Gen Psychiatry 1993; 50: 169–177.
Selemon LD, Rajkowska G, Goldman-Rakic PS . Abnormally high neuronal density in the schizophrenic cortex. A morphometric analysis of prefrontal area 9 and occipital area 17. Arch Gen Psychiatry 1995; 52: 805–818.
Condé F, Lund JS, Jacobowitz DM, Baimbridge KG, Lewis DA . Local circuit neurons immunoreactive for calretinin, calbindin D-28k or parvalbumin in monkey prefrontal cortex: distribution and morphology. J Comp Neurol 1994; 341: 95–116.
Glantz LA, Lewis DA . Decreased dendritic spine density on prefrontal cortical pyramidal neurons in schizophrenia. Arch Gen Psychiatry 2000; 57: 65–73.
Bourgeois JP, Rakic P . Changes of synaptic density in the primary visual cortex of the macaque monkey from fetal to adult stage. J Neurosci 1993; 13: 2801–2820.
Benes FM . Myelination of cortical-hippocampal relays during late adolescence. Schizophr Bull 1989; 15: 585–593.
Lodge DJ . The medial prefrontal and orbitofrontal cortices differentially regulate dopamine system function. Neuropsychopharmacology 2011; 36: 1227–1236.
Selemon LD, Goldman-Rakic PS . The reduced neuropil hypothesis: a circuit based model of schizophrenia. Biol Psychiatry 1999; 45: 17–25.
Jolles DD, van Buchem MA, Crone EA, Rombouts SARB . A comprehensive study of whole-brain functional connectivity in children and young adults. Cereb Cortex 2011; 21: 385–391.
Dauvermann M, Mukherjee P, Moorhead W, Stanfield A, Fusar-Poli P, Lawrie S et al. Relationship between gyrification and functional connectivity of the prefrontal cortex in subjects at high genetic risk of schizophrenia. Curr Pharm Des 2012; 18: 434–442.
Meyer-Lindenberg A . From maps to mechanisms through neuroimaging of schizophrenia. Nature 2010; 468: 194–202.
Kahn RS, Sommer IE . The neurobiology and treatment of first-episode schizophrenia. Mol Psychiatry 2015; 20: 84–97.
Cremer H, Lange R, Christoph A, Plomann M, Vopper G, Roes J et al. Inactivation of the N-CAM gene in mice results in size reduction of the olfactory bulb and deficits in spatial learning. Nature 1994; 367: 455–459.
Wood GK, Tomasiewicz H, Rutishauser U, Magnuson T, Quirion R, Rochford J et al. NCAM-180 knockout mice display increased lateral ventricle size and reduced prepulse inhibition of startle. Neuroreport 1998; 9: 461–466.
Stork O, Welzl H, Cremer H, Schachner M . Increased intermale aggression and neuroendocrine response in mice deficient for the neural cell adhesion molecule (NCAM). Eur J Neurosci 1997; 9: 1117–1125.
Kohl C, Riccio O, Grosse J, Zanoletti O, Fournier C, Klampfl SM et al. The interplay of conditional NCAM-knockout and chronic unpredictable stress leads to increased aggression in mice. Stress 2013; 16: 647–654.
Todaro L, Puricelli L, Gioseffi H, Pallotta MG, Lastiri J, Joffé EBDK et al. Neural cell adhesion molecule in human serum. Increased levels in dementia of the Alzheimer type. Neurobiol Dis 2004; 15: 387–393.
Aisa B, Gil-Bea FJ, Solas M, García-Alloza M, Chen CP, Lai MK et al. Altered NCAM expression associated with the cholinergic system in alzheimer’s disease. J Alzheimer’s Dis 2010; 20: 659–668.
Yew DT, Li WP, Webb SE, Lai HWL, Zhang L . Neurotransmitters, peptides, and neural cell adhesion molecules in the cortices of normal elderly humans and Alzheimer patients: a comparison. Exp Gerontol 1999; 34: 117–133.
Lyons F, Martin ML, Maguire C, Jackson A, Regan CM, Shelley RK . The expression of an N-CAM serum fragment is positively correlated with severity of negative features in type II schizophrenia. Biol Psychiatry 1988; 23: 769–775.
Zhang W, Xiao M-S, Ji S, Tang J, Xu L, Li X et al. Promoter variant rs2301228 on the neural cell adhesion molecule 1 gene confers risk of schizophrenia in Han Chinese. Schizophr Res 2014; 160: 88–96.
Hildebrandt H, Becker C, Glüer S, Rösner H, Gerardy-Schahn R, Rahmann H . Polysialic acid on the neural cell adhesion molecule correlates with expression of polysialyltransferases and promotes neuroblastoma cell growth. Cancer Res 1998; 58: 779–784.
Vawter MP, Cannon-Spoor HE, Hemperly JJ, Hyde TM, VanderPutten DM, Kleinman JE et al. Abnormal expression of cell recognition molecules in schizophrenia. Exp Neurol 1998; 149: 424–432.
Sullivan PF, Keefe RSE, Lange LA, Lange EM, Stroup TS, Lieberman J et al. NCAM1 and neurocognition in schizophrenia. Biol Psychiatry 2007; 61: 902–910.
Bonfanti L . PSA-NCAM in mammalian structural plasticity and neurogenesis. Prog Neurobiol 2006; 80: 129–164.
Frasca A, Fumagalli F, Ter Horst J, Racagni G, Murphy KJ, Olanzapine Riva MA . but not haloperidol, enhances PSA-NCAM immunoreactivity in rat prefrontal cortex. Int J Neuropsychopharmacol 2008; 11: 591–595.
Acknowledgements
We thank Hannelore Burkhardt for expert technical assistance. This paper was funded by the European Commission ERA-Net NEURON joint transnational calI 2010. 'European Research Projects on Mental Disorders': NEUCONNECT. Novel strategies for the treatment of SZ based on genetic variation of the NCAM and enzymes involved in its post-translational modifications. This work was supported by BMBF grant 01EW1106/NeuConnect in the frame of ERA-NET NEURON.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Translational Psychiatry website
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Piras, F., Schiff, M., Chiapponi, C. et al. Brain structure, cognition and negative symptoms in schizophrenia are associated with serum levels of polysialic acid-modified NCAM. Transl Psychiatry 5, e658 (2015). https://doi.org/10.1038/tp.2015.156
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/tp.2015.156
This article is cited by
-
Association of polysialic acid serum levels with schizophrenia spectrum and bipolar disorder-related structural brain changes and hospitalization
Scientific Reports (2023)
-
Ubiquitous Neural Cell Adhesion Molecule (NCAM): Potential Mechanism and Valorisation in Cancer Pathophysiology, Drug Targeting and Molecular Transductions
Molecular Neurobiology (2022)
-
Acute stress-induced change in polysialic acid levels mediated by sialidase in mouse brain
Scientific Reports (2019)
-
Differential effect of disease-associated ST8SIA2 haplotype on cerebral white matter diffusion properties in schizophrenia and healthy controls
Translational Psychiatry (2018)
-
Mental disorders and an acidic glycan-from the perspective of polysialic acid (PSA/polySia) and the synthesizing enzyme, ST8SIA2
Glycoconjugate Journal (2018)
