
Abstract
Some scholars caution that long-term ad libitum feeding with probiotic fermented food poses potential health risks to baby animals. We conducted a feeding experiment to investigate the influence of ad libitum feeding of pre-and post-weaned piglets with a Bacillus subtilis fermented diet on the gut microbiome, gut metabolomic profiles, bile acid metabolism, proinflammatory cytokines and faecal consistency. Compared with piglets fed a Bacillus subtilis-supplemented pellet diet, piglets fed the Bacillus subtilis fermented liquid diet had lower intestinal bacterial diversity (P > 0.05), higher intestinal fungal diversity (P > 0.05), more Firmicutes (P > 0.05), fewer Bacteroidetes, Actinobacteria and Proteobacteria (P > 0.05), higher concentrations of 3-hydroxypropionic acid (P < 0.05), orotic acid (P < 0.05), interleukin-6 (P < 0.01), lactic acid (P < 0.01), deoxycholic acid (P > 0.05) and lithocholic acid (P < 0.01) and a higher incidence of diarrhoea (P > 0.05). The data show that ad libitum feeding of piglets with a Bacillus subtilis fermented liquid diet during the suckling and early post-weaning periods promotes the growth of lactic acid bacteria, bile salt hydrolase-active bacteria and 7a-dehydroxylase-active bacteria in the intestinal lumen; disturbs the normal production of lactic acid, orotic acid and unconjugated bile acids; and increases circulating interleukin-6 levels and diarrhoea incidence.
Similar content being viewed by others


Introduction
Probiotics have been proven to be useful in rebalancing the intestinal flora, improving inflammation and digestion and preventing cardiovascular diseases1,2,3,4, and as a result probiotics are now widely used in food processing and disease control and prevention. To improve the health and growth of children and young animals, specific probiotics are often added to their food at varying doses. Experiments have been conducted to investigate the effects of the routine intake and excessive intake of probiotics on intestinal flora composition, digestion and intestinal health in calves, lambs, piglets and human infants. The resulting data have shown that feeding a moderate dose of probiotics to calves and lambs improves their health and performance5,6,7,8,9. Kukkonen et al. reported thatthe daily feeding of probiotics to newborn human infants for 6 months using an 8–9 × 109 colony-forming-unit mixture of specific probiotics was safe10. However, others have argued that supplementing the daily food of infants with probiotics should be done with caution or not at all11,12,13 because of the underdeveloped state of the infant immune system14. Li et al. (2012) found that oral administration of Lactobacillus rhamnosus at a high dose to piglets resulted in severe diarrhoea15. Thus, there is still controversy regarding the safety and impact of probiotics on young animals, particularly regarding the strains, dosage and duration of probiotic administration. These factors should be taken into account as different strains, dosages and durations may have drastically different effects than intended16. More information is needed regarding the long-term safety of probiotics and probiotic fermented food11,13, especially regarding lactic acidosis and bile salt malabsorption caused by bacterial overgrowth; these issues have been rarely studied11.
The intestinal flora of pigs plays important roles in intestinal morphology, immunity, digestion and health17,18,19. Generally, from a phylum-level perspective, the flora of the pig intestine can be classified into five phyla: Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria and Spirochaetes. Firmicutes represents the largest proportion of the total population, followed by Bacteroidetes. These two phyla account for approximately 90% of all the bacteria present in the pig intestine. However, the intestinal microbiota is dynamic, and its composition changes continually in response to time, age, diet, probiotics and many other factors20.
In the present study, suckling piglets were used as a model to study the influence of long-term ad libitum feeding of a Bacillus subtilis fermented liquid diet on intestinal flora composition, pH, unconjugated bile acids, inflammation and diarrhoea in order to inform risk assessments and investigate the safety of using Bacillus subtilis fermented products as a daily food for baby monogastric animals.
Results
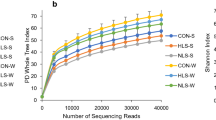
Operational Taxonomic Unit (OTU) and Alpha Diversity
The sequence data produced in this experiment have been deposited in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) under accession number SRP060218. Data on OTU and alpha diversity of the microorganism communities in different dietary treatment groups are listed in Table 1 The OTU number, Chao 1 and Shannon values of bacterial communities in the jejunal luminal content of weaned piglets from the Bacillus subtilis fermented liquid diet (BFLD) group were lower (P > 0.05) than those of weaned piglets from the Bacillus subtilis-supplemented commercial pellet diet (BCPD) group. In contrast, the OTU numbers and Chao 1 values of bacterial communities in the colonic luminal content of weaned piglets from the BFLD group were higher (P > 0.05) than those of weaned piglets from the BCPD group, which did not hold true for the Shannon value. These results indicated that the feeding of Bacillus subtilis fermented liquid diet to piglets decreased bacterial richness and diversity in the jejunal luminal content and decreased bacterial diversity in the colonic luminal content. The Chao 1 values of the fungal community were lower (P > 0.05) in the jejunal luminal content but were higher (P > 0.05) in the colonic luminal content of weaned piglets from the BFLD group than those of weaned piglets from the BCPD group. These findings suggest that feeding with a Bacillus subtilis fermented liquid diet increases fungal diversity in the jejunal and colonic luminal contents of weaned piglets compared with feeding with a Bacillus subtilis-supplemented pellet diet.
Compositions and Relative Abundances of Microorganisms in Jejunal and Colonic Luminal Contents
The compositions and relative abundances of microorganisms in the jejunal and colonic luminal contents are shown in Tables 2 and 3, respectively. Firmicutes and Ascomycota were the dominant phyla in the jejunal and colonic luminal contents of weaned piglets fed either the Bacillus subtilis fermented liquid diet or the Bacillus subtilis-supplemented commercial pellet diet. Compared with weaned piglets from the BCPD group, weaned piglets from the BFLD group had a higher (P > 0.05) relative abundance of Firmicutes in the jejunal and colonic luminal contents, a lower (P > 0.05) relative abundance of Ascomycota in the jejunal luminal content and a higher (P > 0.05) relative abundance of Ascomycota in the colonic luminal content. The relative abundances of Bacteroidetes, Actinobacteria and Proteobacteria were lower (P > 0.05) in the jejunal and colonic luminal contents of weaned piglets from the BFLD group compared with those from the BCPD group. At the genus level, Lactobacillus and Kazachstania were the dominant genera in jejunal and colonic luminal contents of weaned piglets from both the BFLD and BCPD groups. The relative abundance of Lactobacillus in the jejunal and colonic luminal contents of weaned piglets from the BFLD group was higher (P > 0.05) than that of weaned piglets from the BCPD group. Weaned piglets from the BFLD group had a lower (P > 0.05) relative abundance of Kazachstania in the jejunal luminal content and a higher (P > 0.05) relative abundance of Kazachstania in the colonic luminal content compared with weaned piglets from the BCPD group. The relative abundances of Streptococcus, Clostridium_sensu_stricto, Bacteroides and Flavobacterium in the jejunal luminal content of weaned piglets from the BFLD group were significantly lower (P < 0.01 or P < 0.05) than those of weaned piglets from the BCPD group. The relative abundances of Pseudobutyrivibrio, Lachnospiraceae_unclassified, Erysipelotrichaceae_unclassified, Ruminococcus, Clostridiales_unclassified and Lachnospiraceae_uncultured in the colons of weaned piglets from the BFLD group were significantly higher (P < 0.01 or P < 0.05) than those of weaned piglets from the BCPD group.
Differential Metabolite Levels in Jejunal and Colonic Luminal Contents
All metabolites found at levels that differed between the two piglet groups are listed in Tables 4 and 5. Thirteen differentially observed metabolites in the jejunal luminal content and eleven differentially observed metabolites in the colonic luminal content were identified. Piglets from the BFLD group had higher (P < 0.05) relative levels of 3-hydroxypropionic acid and orotic acid in their jejunal luminal content and higher (P < 0.05) relative levels of stigmasterol in their colonic luminal content than piglets from the BCPD group. Except for 3-hydroxypropionic acid, orotic acid and stigmasterol, the relative levels of other differentially observed metabolites in the jejunal and colonic luminal contents of weaned piglets from the BFLD group were significantly lower (P < 0.01 or P < 0.05) than those of piglets from the BCPD group.
Serum Cytokines, Intestinal pH and Unconjugated Bile Acids
Piglets from the BFLD group had significantly higher serum interleukin-6 (IL-6) levels (P < 0.01) than piglets from the BCPD group (Table 6). There were no significant differences (P > 0.05) in the levels of serum tumour necrosis factor-alpha (TNF-α) and IL-1β between the BFLD and BCPD groups.
There was no significant difference in the pH values of the jejunal luminal content between the BFLD and BCPD groups (P > 0.05). However, the pH values of the colonic luminal content collected from piglets in the BFLD group were significantly lower than the pH values of the colonic luminal content collected from piglets in the BCPD group (P < 0.05).
The jejunal luminal content collected from the BFLD group had significantly higher lactic acid and lithocholic acid (LCA) concentrations (P < 0.01) and significantly lower cholic acid (CA) concentrations (P < 0.05) than that collected from the BFLD group. There were no significant differences in chenodeoxycholic acid (CDCA) and deoxycholic acid (DCA) concentrations in the jejunal luminal content between the BFLD and BCPD groups (P > 0.05).
The colonic luminal content collected from the BFLD group had significantly higher lactic acid, chenodeoxycholic acid and lithocholic acid concentrations than that collected from the BCPD group (P < 0.01). The concentrations of cholic acid and deoxycholic acid in the colonic luminal content collected from the BFLD group were not significantly higher than those in the colonic content collected from the BCPD group (P > 0.05).
Diarrhoea Incidence
The data in Table 7 show that piglets from the BFLD group had a higher incidence of diarrhoea than piglets from the BCPD group at each experimental time point, but there was no significant difference (P > 0.05) in the incidence of diarrhoea between the BFLD and BCPD groups.
Discussion
Previous studies have reported that the diversity, composition and relative abundance of intestinal flora can be influenced by probiotic administration or dietary patterns21,22. The feeding of probiotics and a probiotic fermented diet to animals decreases microbial diversity, and the reduced microbial diversity is often associated with gastrointestinal disorders, including inflammatory bowel disease23. In the present study, 25-day feeding with a Bacillus subtilis fermented liquid diet to piglets aged 7 to 31 days decreased the bacterial diversity but increased the fungal diversity of jejunal and colonic luminal contents compared with 25-day feeding with a Bacillus subtilis-supplemented pellet diet; the decreased bacterial diversity of piglets fed a Bacillus subtilis fermented liquid diet resulted in a higher diarrhoea incidence than observed in the piglets fed a Bacillus subtilis-supplemented pellet diet. In addition, Ley et al. reported that the gut microbiome is dominated by four bacterial phyla: Firmicutes, Bacteroidetes, Actinobacteria and Proteobacteria24; data in this study also indicated that the flora in the jejunal and colonic luminal contents of piglets fed continuously with a Bacillus subtilis fermented liquid diet or a Bacillus subtilis-supplemented pellet diet were also dominated by Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria and Ascomycota. However, different dietary patterns influenced the relative abundances of intestinal flora: piglets fed a Bacillus subtilis fermented liquid diet had higher (P > 0.05) relative abundances of organisms from the Firmicutes phylum and Lactobacillus genus in their jejunal luminal contents than piglets fed a Bacillus subtilis-supplemented pellet diet. Regarding the relative abundances of flora in the colonic luminal contents, piglets fed a Bacillus subtilis fermented liquid diet not only had higher (P > 0.05) relative abundances of organisms of the Firmicutes phylum and Lactobacillus genus but also had higher (P > 0.05) relative abundances of organisms of the Ascomycota phylum and Kazachstania genus than did piglets fed a Bacillus subtilis-supplemented pellet diet. These results indicate that a Bacillus subtilis fermented diet has an advantage in promoting the growth of the flora noted above because the Bacillus subtilis fermented liquid diet has ingredients more suitable for the growth of intestinal flora than the Bacillus subtilis-supplemented pellet diet.
Orotic acid is often regarded as one of the major oxidative stressors at high concentrations25. Additionally, circulating levels of IL-6 and the growth of Coprococcus, Pseudobutyrivibrio and Dorea increase under the action of stressors26. Piglets from the BFLD group had significantly higher orotic acid levels in their jejunal luminal content than piglets from the BCPD group. As a result, piglets from the BFLD group had higher circulating IL-6 levels and higher relative abundances of Coprococcus, Pseudobutyrivibrio and Dorea than piglets from the BCPD group.
Elevated levels of circulating IL-6 are often associated with a number of diseases27. People with high IL-6 levels have a high risk of systemic mastocytosis28, and elevated circulating IL-6 has been proposed as a marker of inflammation linking obesity with insulin resistance and diabetes as well as atherosclerosis29,30. High serum levels of IL-6 may also be associated with ankylosing spondylitis in young people, which is characterized by intense joint pain, stiffness, weakness, marasmus and apocleisis31.
Studies have demonstrated that diarrhoea can be prevented by the administration of probiotics or probiotic fermented food32. However, Li et al. (2012) found that oral administration of Lactobacillus rhamnosus at a high dose to piglets caused diarrhoea15. Data in the present study also showed that piglets from the BFLD group had a higher incidence of diarrhoea than piglets from the BCPD group.
Conjugated bile acids have emulsifying and surfactant properties; they are more efficient than unconjugated bile acids in aiding in the emulsification of dietary lipids and preventing small intestinal bacterial overgrowth33,34. In normal conditions, the composition of bile acids in the intestine is often in a relative balance; only small amounts of conjugated bile acids are hydrolyzed into primary bile acids (CA and CDCA), and small amounts of primary bile acids are dehydroxylated into secondary bile acids (DCA and LCA) in the small intestine35. Approximately 95% of bile acids (conjugated and unconjugated) are reabsorbed by the distal ileum. The small percentage of bile acids remaining reaches the colon, where they are deconjugated and dehydroxylated by bacteria to produce the secondary bile acids (DCA and LCA)36. However, overgrowth of bile salt hydrolase-active and 7a-dehydroxylase-active bacteria in the intestine will alter the normal bile acid composition and damage normal enterohepatic circulation.
Some strains of Lactobacillus37, Erysipelotrichaceae38, Lachnospiraceae39, Clostridium40,41 and Bacteroides42,43 are bile salt hydrolase-active intestinal bacteria, and some strains of Lactobacillus44, Lachnospiraceae45,46, Ruminococcaceae45, Clostridiaceae46,47, Eubacterium48 and Peptostreptococcus49 are 7a-dehydroxylase-active intestinal bacteria. In the present study, compared with piglets from the BCPD group, piglets from the BFLD group had lower (P < 0.01) relative abundances of Clostridium and Bacteroides in their jejunal luminal content. This condition resulted in piglets from the BFLD group having lower CA and CDCA levels in their jejunal luminal content. Lactobacilli are also 7a-dehydroxylase-active bacteria, and the relative abundance of Lactobacilli in the jejunal luminal content of piglets from the BFLD group were higher than those in piglets from the BCPD group. This is the reason why piglets from the BFLD group had higher DCA and LCA levels in their jejunal luminal content than piglets from the BCPD group did. Piglets from the BFLD group also had higher relative abundances of Lactobacillus, Ruminococcaceae, Lachnospiraceae, Erysipelotrichaceae, Ruminococcus and Clostridiales in their colonic luminal contents than piglets from the BCPD group. Therefore, in the BFLD piglets, more conjugated bile acids were hydrolyzed into CA and CDCA, while CA and CDCA were dehydroxylated into DCA and LCA, respectively. As a result, piglets from the BFLD group had higher CA, CDCA, DCA and LCA levels in their colonic luminal content than piglets from the BCPD group.
Unconjugated bile acids are less water soluble than conjugated bile acids. Intense elevation of the concentration of unconjugated bile acids has detrimental effects on the intestinal mucosa, including mucosal damage, increased mucosal permeability and potentially colon cancer-promoting effects50,51,52. Increased CDCA and DCA levels can inhibit water absorption and induce water and sodium secretion by the colon at concentrations above 3 mmol/L53 and can disturb the normal microbiota of the gut, leading to diarrhoea and mucosal inflammation in the intestinal contents54. Piglets from the BFLD group had higher CDCA and DCA in their colonic luminal contents than piglets from the BCPD group; this is one of the factors contributing to the higher diarrhoea incidence among piglets from the BFLD group.
The Bacillus subtilis fermented liquid diet contains more active components for the growth of lactic acid bacteria and a high lactic acid concentration55,56. These active components allowed lactic acid-producing bacteria to grow better in the intestines of piglets from the BFLD group than in the intestines of piglets from the BCPD group. The higher relative abundance of lactic acid bacteria together with the high lactic acid intake resulted in piglets from the BFLD group having higher lactic acid in their jejunal and colonic luminal contents than piglets from the BCPD group. Excessive lactic acid in the intestine often causes lactic acidosis, which can induce diarrhoea57. Thus, lactic acidosis is the other factor contributing to the higher diarrhoea incidence among piglets from the BFLD group than among piglets from the BCPD group.
In summary, ad libitum feeding of pre-and post-weaned piglets with a Bacillus subtilis fermented liquid diet decreased intestinal bacterial diversity and increased intestinal fungal diversity, circulating IL-6 levels, intestinal unconjugated bile acid concentrations and diarrhoea incidence. Lactic acidosis, dietary lipid malabsorption and the inducing effect of unconjugated bile salts are the underlying causes for the higher diarrhoea incidence among piglets fed the Bacillus subtilis fermented liquid diet.
Materials and Methods
Animal Treatment Protocol
Twelve lactating sows (Large White x Landrace, artificially inseminated with semen from one Duroc boar) with similar body conditions and suckling piglets were randomly assigned to one experimental group and one control group (6 lactating sows +56 suckling piglets vs 6 lactating sows +54 suckling piglets) at the 7th day after farrowing. There was no significant difference (P > 0.05) in the average body weight (2.79 ± 0.19 kg/piglet vs 2.85 ± 0.27 kg/piglet) of suckling piglets at 7 days of age between the experimental and control groups. All lactating sows were fed the same commercial lactation diet (7.5 kg/d). Piglets in the experimental group and control group had free access to a Bacillus subtilis fermented liquid diet (live Bacillus subtilis: 12.75 × 108 CFU/g) or a Bacillus subtilis-supplemented commercial pellet diet (live Bacillus subtilis: 2.80 × 108 CFU/g), respectively, from 7 to 31 days of age. All suckling piglets were weaned at 21 days of age. The Bacillus subtilis fermented liquid diet was produced using a previously described method46.
A total of six piglets (each with a body weight closest to the average weight of the litter) in each dietary treatment group (3 males and 3 females) were slaughtered in the morning at 32 days of age according to the protocol approved by the Animal Ethics Committee of Jiangxi Agricultural University.
Sample Collection
Before slaughter, blood was collected with a 10 mL fresh tube from the jugular vein. Serum was separated by centrifugation after blood clotting and stored at −20 °C for the analysis of serum cytokines.
After slaughter, the segments of the jejunum and colon were quickly excised. The contents of the jejunum and colon were separately collected with 10 mL fresh tubes and immediately stored at −80 °C for the analysis of pH, lactic acid, unconjugated bile acids, microbial composition and differential metabolites.
pH Measurement
A digital pH metre (LP115FK, Mettler Toledo, Switzerland) was used to measure the pH of samples after calibration with standard buffers (pH 4.0 and 7.0).
Enzyme-linked Immunosorbent Assay
The concentrations of interleukin −1β, interleukin −6 and tumour necrosis factor-alpha were determined in three replicates for each sample using enzyme-linked immunosorbent assay kits (R&D Systems, Nanjing Jiancheng Bioengineering Institute).
Concentrations of D-/L-lactic acid, cholic acid, chenodeoxycholic acid, deoxycholic acid and lithocholic acid in the intestinal contents were determined in three replicates for each sample using enzyme-linked immunosorbent assay kits (R&D Systems, Shanghai Enzyme-linked Biotechnology Co., Ltd.).
Microbiome and Metabolomics Analysis
Genomic DNA of each sample was extracted using the E.Z.N.A. Soil DNA kit (OMEGA, USA), and six genomic DNA preparations for each treatment group were pooled into three DNA mixtures prior to polymerase chain reaction (PCR).
Bacterial genomic DNA was amplified with primers covering the V1–V3 region of the 16 S rRNA bacterial gene; the bar-coded primers 27 F and 533 R containing A and B sequencing adaptors (454 Life Sciences) were used. The forward primer (B-27F) was 5′-CCTATCCCCTGTGTGCCTTGGCAGTCGACTAGAGTTTGATCCTGGCTCAG-3′; the sequence of the B adaptor is shown in italics and is underlined. The reverse primer (A-533R) was 5′-CCATCTCATCCCTGCGTGTCTCCGACGACTNNNNNNNNNNTTACCGCGGCTGCTGGCAC-3′; the sequence of the A adaptor is shown in italics and underlined, and the Ns represent a ten-base sample specific barcode sequence58.
Fungal genomic DNA was amplified using the forward primer (A-ITS1) and reverse primer (B-ITS4). The forward primer (A-ITS1) was 5′-CCATCTCATCCCTGCGTGTCTCCGACGACTNNNNNNNNNNTCCGTAGGTGAACCTGCGG-3′; the sequence of adaptor A is shown in italics and underlined, and the Ns represent a ten-base sample specific barcode sequence. The reverse primer (B-ITS4) was 5′-CCTATCCCCTGTGTGCCTTGGCAGTCGACTTCCTCCGCTTATTGATATGC-3′, and the sequence of adaptor B is shown in italics and underlined.
The protocols for PCR, pyrosequencing, sequence processing and bioinformatic analyses were previously described56,59. Differential metabolites were determined using gas chromatography time of flight mass spectrometry55.
Diarrhoea Incidence Calculation
Faecal consistency was visually examined at the same time each morning by the same person during experimental periods. A piglet was considered to have diarrhoea when it developed a pasty or watery faecal consistency. Diarrhoea incidence was defined as the percentage of animals with diarrhoea on a specific day60.
Statistical Analysis
Data analysis was performed with SPSS software (version 13.0). One-way analysis of variance (ANOVA) was used to evaluate significant differences between means with a significance level at α = 0.01. Tukey’s test was used to perform comparisons between means. Data were presented as the means ± SEM.
Additional Information
How to cite this article: He, Y. et al. Influence of ad libitum feeding of piglets with Bacillus subtilis fermented liquid feed on gut flora, luminal contents and health. Sci. Rep. 7, 44553; doi: 10.1038/srep44553 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Marques-Lopes, I., Ansorena, D., Astiasaran, I., Forga, L. & Martínez, J. A. Postprandial de novo lipogenesis and metabolic changes induced by a high-carbohydrate, low-fat meal in lean and overweight men. Am J Clin Nutr 73, 253–261 (2001).
Riggs, A. J., White, B. D. & Gropper, S. S. Changes in energy expenditure associated with ingestion of high protein, high fat versus high protein, low fat meals among underweight, normal weight, and overweight females. Nutrition J. 6, 40, doi: 10.1186/1475-2891-6-40 (2007).
Kalil, A. C. & Schooneveld, T. C. Probiotics and antibiotic-associated diarrhoea. Lancet 383, 29–30, doi: 10.1016/S0140-6736(13)62734-8 (2014).
Ford, A. C. et al. Efficacy of prebiotics, probiotics, and synbiotics in irritable bowel syndrome and chronic idiopathic constipation: systematic review and meta-analysis. Am J Gastroenterol. 109, 1547–1561, doi: 10.1038/ajg.2014.202 (2014).
Agazzi, A., Tirloni, E., Stella, S. & Savoini, G. Effects of species-specific probiotic addition to milk replacer on calf health and performance during the first month of life. Ann Anim Sci. 14, 101–115, doi: 10.2478/aoas-2013-0089 (2014).
Noori, M., Alikhani, M. & Jahanian, R. Effect of partial substitution of milk with probiotic yogurt of different pH on performance, body conformation and blood biochemical parameters of Holstein calves. J Appl Anim Res. 44, 1–9, doi: 10.1080/09712119.2015.1031772 (2015).
Timmerman, H. M. et al. Health and Growth of Veal Calves Fed Milk Replacers With or Without Probiotics. J Dairy Sci. 88, 2154–2165, doi: 10.3168/jds.S0022-0302(05)72891-5 (2005).
Kowalski, Z. M. et al. Performance of Holstein calves fed milk-replacer and starter mixture supplemented with probiotic feed additive. J Anim Feed Sci. 18, 399–411 (2009).
Santillo, A. et al. Probiotics in milk replacer influence lamb immune function and meat quality. Animal 6, 339–345, doi: 10.1017/S1751731111001571 (2012).
Kukkonen, K. et al. Long-term safety and impact on infection rates of postnatal probiotic and prebiotic (synbiotic) treatment: randomized, double-blind, placebo-controlled trial. Pediatrics 122, 8–12, doi: 10.1542/peds.2007-1192 (2008).
Braegger, C. et al. Supplementation of infant formula with probiotics and/or prebiotics: a systematic review and comment by the ESPGHAN committee on nutrition. J Pediatr Gastr Nutr. 52, 238–250, doi: 10.1097/MPG.0b013e3181fb9e80 (2011).
Sarkar, S. Probiotics, Prebiotics and Synbiotics for Infant Feeding – A Review. J Microbial Biochem Technol S1, 004, doi: g/10.4172/1948-5948.S1-004 (2011).
Shafai, T. Routine supplement of prebiotics and probiotics to newborn infants is not recommended. Pediatrics 123, e543–544, doi: 10.1542/peds.2008-3557 (2009).
Kobyliak, N. et al. Probiotics in prevention and treatment of obesity: a critical view. Nutr Metab. 13, 14, doi: 10.1186/s12986-016-0067-0 (2016).
Li, X. Q. et al. Risks associated with high-dose Lactobacillus rhamnosus in an Escherichia coli model of piglet diarrhoea: intestinal microbiota and immune imbalances. PLoS One 7, e40666, doi: 10.1371/journal.pone.0040666 (2012).
Ritchie, M. L. & Romanuk, T. N. A meta-analysis of probiotic efficacy for gastrointestinal diseases. Plos One 7, e34938, doi: 10.1371/journal.pone.0034938 (2012).
Niu, Q. et al. Dynamic distribution of the gut microbiota and the relationship with apparent crude fiber digestibility and growth stages in pigs. Sci Rep 5, 9938, doi: 10.1038/srep09938 (2015).
Ashida, H., Ogawa, M., Kim, M., Mimuro, H. & Sasakawa, C. Bacteria and host interactions in the gut epithelial barrier. Nat Chem Biol 8, 36–45 (2012).
Kim, H. B. et al. Microbial shifts in the swine distal gut in response to the treatment with antimicrobial growth promoter, tylosin. Proc Natl Acad Sci USA 109, 15485–15490 (2012).
Isaacson, R. & Kim, H. B. The intestinal microbiome of the pig. Anim Health Res Rev 13, 100–109 (2012).
Goossens, D. A., Jonkers, D. M., Russel, M. G., Stobberingh, E. E. & Stockbrügger, R. W. The effect of a probiotic drink with Lactobacillus plantarum 299 v on the bacterial composition in faeces and mucosal biopsies of rectum and ascending colon. Aliment Pharmacol Ther. 23, 255–263, doi: 10.1111/j.1365-2036.2006.02749.x (2006).
Delzenne, N. M., Neyrinck, A. M., Bäckhed, F. & Cani, P. D. Targeting gut microbiota in obesity: effects of prebiotics and probiotics. Nat Rev Endocrinol. 7, 639–646, doi: 10.1038/nrendo.2011.126 (2011).
Ott, S. J. et al. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut 53, 685–693 (2004).
Ley, R. E., Knight, R. & Gordon, J. I. The human microbiome: eliminating the biomedical/environmental dichotomy in microbial ecology. Environ Microbiol. 9, 3–4, doi: 10.1111/j.1462-2920.2006.01222_3.x (2007).
Konstantinov, S. R. et al. S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. PNAS 105, 19474–19479, doi: 10.1073/pnas.0810305105 (2008).
Bailey, M. T. et al. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav Immun. 25, 397–407, doi: 10.1016/j.bbi.2010.10.023 (2011).
Heinrich, P. C., Castell, J. V. & Andus, T. Interleukin-6 and the acute phase response. Biochem J 265, 621–636 (1990).
Mayado, A. et al. Increased IL6 plasma levels in indolent systemic mastocytosis patients are associated with high risk of disease progression. Leukemia 30, 124–130 (2016).
Eder, K., Baffy N. & Falus A. The major inflammatory mediator interleukin-6 and obesity. Inflamm Res 58, 727–736, doi: 10.1007/s00011-009-0060-4 (2009).
Hosick, P. A. et al. Differences in the GH-IGF-I axis in children of different weight and fitness status. Growth Horm IGF Res 22, 87–91, doi: 10.1016/j.ghir.2012.02.007 (2012).
Liu, W. et al. Elevated serum levels of IL-6 and IL-17 may associate with the development of ankylosing spondylitis. Int J Clin Exp Med 8, 17362–17376 (2015).
Sánchez, B. et al. Probiotics, gut microbiota, and their influence on host health and disease. Mol Nutr Food Res. doi: 10.1002/mnfr.201600240 (2016).
Lorenzo-Zúñiga, V. et al. Oral bile acids reduce bacterial overgrowth, bacterial translocation, and endotoxemia in cirrhotic rats. Hepatology 37, 551–557, doi: 10.1053/jhep.2003.50116 (2003).
Hylemon, P. B. et al. Bile acids as regulatory molecules. J Lipid Res. 50, 1509–1520, doi: 10.1194/jlr.R900007-JLR200 (2009).
Zhang, Y., Limaye, P. B., Renaud, H. J. & Klaassen, C. D. Effect of various antibiotics on modulation of intestinal microbiota and bile acid profile in mice. Toxicol Appl Pharmacol 277, 138–145, doi: 10.1016/j.taap.2014.03.009 (2014).
Hofmann, A. F. & Hagey, L. R. Bile acids: Chemistry, pathochemistry, biology, pathobiology, and therapeutics. Cell Mol Life Sci. 65, 2461–2483, doi: 10.1007/s00018-008-7568-6 (2008).
Moser, S. A. & Savage, D. C. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in lactobacilli. Appl Environ Microbiol. 67, 3476–3480, doi: 10.1128/AEM.67.8.3476-3480.2001 (2001).
Labbé, A., Ganopolsky, J. G., Martoni, C. J., Prakash, S. & Jones, M. L. Bacterial bile metabolising gene abundance in Crohn’s, ulcerative colitis and type 2 diabetes metagenomes. PLoS One 9, e115175, doi: 10.1371/journal.pone.0115175 (2014).
Stevenson, B. S., Eichorst, S. A., Wertz, J. T., Schmidt, T. M. & Breznak, J. A. New strategies for cultivation and detection of previously uncultured microbes. Appl Environ Microb. 70, 4748–4755, doi: 10.1128/AEM.70.8.4748-4755.2004 (2004).
Coleman, J. P. & Hudson, L. L. Cloning and characterization of a conjugated bile acid hydrolase gene from Clostridium perfringens. Appl Environ Microbiol. 61, 2514–2520 (1955).
Pereira, D. I., McCartney, A. L. & Gibson, G. R. An in vitro study of the probiotic potential of a bile-salt-hydrolyzing Lactobacillus fermentum strain, and determination of its cholesterol-lowering properties. Appl Environ Microbiol. 69, 4743–4752 (2003).
Kawamoto, K., Horibe, I. & Uchida, K. Purification and characterization of a new hydrolase for conjugated bile acids, chenodeoxy-cholyltaurine hydrolase, from Bacteroides vulgatus. J Biochem. 106, 1049–1053 (1989).
Stellwag, E. J. & Hylemon, P. B. Purification and characterization of bile salt hydrolase from Bacteroides fragilis subsp. Fragilis. Biochim Bio-phys Acta 452, 165–176 (1976).
Hayakawa, S. Microbiological transformation of bile acids. Adv Lipid Res. 11, 143–192 (1973).
Kakiyama, G. et al. Modulation of the fecal bile acid profile by gut microbiota in cirrhosis. J Hepatol. 58, 949–955, doi: 10.1016/j.jhep.2013.01.003 (2013).
Stellwag, E. J. & Hylemon, P. B. 7a-Dehydroxy-lation of cholic acid and chenodeoxycholic acid by Clostridium leptum. J Lipid Res. 20, 325–333 (1979).
Hirano, S., Nakama, R., Tamaki, M., Masuda, N. & Oda, H. Isolation and characterization of thirteen intestinal microorganisms capable of 7cx-dehydroxylating bile acids. Appi Environ Microbiol. 41, 737–745 (1981).
Lipsky, R. H. & Hylemon, P. B. Characterization of a NADH:flavin oxidoreductase induced by cholic acid in a 7cx-dehydroxylating intestinal Eubacterium species. Biochim Biophys Acta. 612, 328–336 (1980).
Edenharder, R., Pfutzner, A. & Hammann, R. Characterization of NAD-dependent 3 alpha- and 3 beta-hydroxysteroid dehydroge-nase and of NADP-dependent 7 beta-hydroxysteroid dehydro-genase from Peptostreptococcus productus. Biochim Biophys Acta. 1004, 230–238 (1989a).
Marteau, P. et al. Metabolism of bile salts by alimentary bacteria during transit in the human small intestine. Microb Ecol Health Dis. 8, 151–157 (1995).
Nagengast, F. M., Grobben, M. J. & van Munster, I. P. Role of bile acids in colorectal carcinogenesis. Eur J Cancer. 31, 1067–1070 (1995).
Hamer, H. M., De Preter, V., Windey, K. & Verbeke, K. Functional analysis of colonic bacterial metabolism: relevant to health? Am J Physiol Gastrointest Liver Physiol. 302, G1–G9, doi: 10.1152/ajpgi.00048.2011 (2012).
McJunkin, B., Fromm, H., Sarva, P. R. & Amin, P. Factors in the mechanism of diarrhea in bile acid malabsorption:fecal pH-a key determinant. Gastroenterology 80, 1454–1464 (1981).
Salminen, S., Isolauri, E. & Salminen, E. Clinical uses of probiotics for stabilizing the gut mucosal barrier: Successful strains for future challenges. Antonie van Leeuwenhoek 70, 347–358 (1996).
He, Y. Y. et al. Identification of differential metabolites in liquid diet fermented with Bacillus subtilis using gas chromatography time of flight mass spectrometry. Anim Nutr. 2, 351–356, doi: 10.1016/j.aninu.2016.07.007 (2016).
He, Y. Y., Chen, Z. Y., Liu, X. L., Wang, C. W. & Lu, W. Influence of trace elements mixture on bacterial diversity and fermentation characteristics of liquid diet fermented with probiotics under air-tight condition. PLos One 9, e114218, doi: 10.1371/journal.pone.0114218 (2014).
Omole, O. O., Nappert, G., Naylor, J. M. & Zello, G. A. Both L- and D-lactate contribute to metabolic acidosis in diarrheic calves. J Nutr. 131, 2128–2131 (2001).
Watanabe, K., Nagao, N., Toda, T. & Kurosawa, N. The dominant bacteria shifted from the order ‘Lactobacillales’ to Bacillales and Actinomycetales during a start-up period of large-scale, completelymixed composting reactor using plastic bottle flakes as bulking agent. World J Microb Biot. 25, 803–811, doi: 10.1007/s11274-008-9952-7 (2009).
He, Y. Y., Chen, Z. Y., Wen, H., Lu, W. & Wu, H. D. Pyrosequencing investigation into the influence of Cu2+, Zn2+, Fe2+ and I- mixtures on fungal diversity and toxigenic fungal growth in a fermented liquid feed. Anim Nutr. 2, 51–56, doi: 10.1016/j.aninu.2016.02.004 (2016).
Kiers, J. L. et al. Effect of fermented soya beans on diarrhoea and feed efficiency in weaned piglets. J Appl Microbiol. 95, 545–552 (2003).
Acknowledgements
The study was funded by the Jiangxi Provincial Key Technology R&D Program (20121BBF60032 and 20132BBF60039).
Author information
Authors and Affiliations
Contributions
Y.H., W.L. and H.W. conceived and designed this study and wrote the manuscript. C.M., Z.C. and H.W. conducted this experiment. H.W., C.M., T.L. and L.L. reviewed the literature and analysed samples.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
He, Y., Mao, C., Wen, H. et al. Influence of ad Libitum Feeding of Piglets With Bacillus Subtilis Fermented Liquid Feed on Gut Flora, Luminal Contents and Health. Sci Rep 7, 44553 (2017). https://doi.org/10.1038/srep44553
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep44553
This article is cited by
-
Lactobacillus plantarum A3 attenuates ulcerative colitis by modulating gut microbiota and metabolism
Animal Diseases (2023)
-
Engineered butyrate-producing bacteria prevents high fat diet-induced obesity in mice
Microbial Cell Factories (2020)
-
Two antimicrobial genes from Aegilops tauschii Cosson identified by the Bacillus subtilis expression system
Scientific Reports (2020)
-
Microbiota in fermented feed and swine gut
Applied Microbiology and Biotechnology (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
