
Abstract
Serotonin (5-HT) is an important neurotransmitter and neuromodulator that controls a variety of sensory and motor functions through 5-HT receptors (5-HTRs). The 5-HT4R subfamily is linked to Gs proteins, which activate adenylyl cyclases (ACs), and is involved in many responses in peripheral organs. In this study, the 5-HT4R from Apostichopus japonicus (Aj5-HT4R) was identified and characterised. The cloned full-length Aj5-HT4R cDNA is 1,544 bp long and contains an open reading frame 1,011 bp in length encoding 336 amino acid proteins. Bioinformatics analysis of the Aj5-HT4R protein indicated this receptor was a member of class A G protein coupled receptor (GPCR) family. Further experiments using Aj5-HT4R-transfected HEK293 cells demonstrated that treatment with 5-HT triggered a significant increase in intracellular cAMP level in a dose-dependent manner and induced a rapid internalisation of Aj5-HT4R fused with enhanced green fluorescent protein (Aj5-HT4R-EGFP) from the cell surface into the cytoplasm. In addition, the transcriptional profiles of Aj5-HT4R in aestivating A. japonicas and phosphofructokinase (AjPFK) in 5-HT administrated A. japonicus have been analysed by real-time PCR assays. Results have led to a basic understanding of Aj5-HT4R in A. japonicus, and provide a foundation for further exploration of the cell signaling and regulatory functions of this receptor.
Similar content being viewed by others

Introduction
The biogenic amine serotonin (5-hydroxytryptamine; 5-HT), first isolated in 19481, is a well-known neurotransmitter and neuromodulator. It has been described by Dr Barnes as the happy hormone2, involved in natural reward-related physiology and behaviour3, from feeding to sexual activity. Serotonin is biochemically derived from tryptophan and has various functions in different phyla, including both vertebrates and invertebrates, and plays a modulatory role in numerous physiological processes such as feed intake, reproduction, immunity and stress responses4,5,6.
Presently, seven classes of 5-HT receptors, 5-HT1 to 5-HT7, have been identified. Aside from 5-HT3, which is a ligand-gated ion channel that belongs to the nicotinic acetylcholine receptor superfamily7, the 5-HT receptors belong to the superfamily of G protein coupled receptors (GPCRs)8,9. Orthologous G protein-coupled 5-HT receptors have been identified and functionally characterised in deuterostomes including humans10, mammals11, birds12 and fishes13, and in protostomes including nematodes14, crustaceans15 and insects16,17. Furthermore, these G protein-coupled 5-HT receptors are mainly classified in three groups: (1) 5-HT1 and 5-HT5 receptors that couple preferentially to Gi proteins and inhibit cAMP synthesis, (2) 5-HT2 receptors that activate Gq proteins, which mediate the hydrolysis of inositol phosphates and cause a subsequent increase in cytosolic Ca2+, and (3) 5-HT4, 5-HT6, and 5-HT7 receptors that couple to Gs proteins and promote cAMP formation18,19. Additionally, an extra novel serotonin receptor (named Pr5-HT8) has been identified from larval Pieris rapae and, and represents the first recognized member of a novel 5-HT receptor class with a unique pharmacological profile20. In marine invertebrates or dormancy animals, only several serotonin receptors have been identified from the barnacle, Balanus amphitrite Darwin21 and the marine mollusc Aplysia22,23, however, the 5-HT receptors and their physiological functions in marine invertebrates still remain largely unknown. Meanwhile, aside from several gene expression analyses on 5-HT receptor-like genes24, our understanding of the molecular and pharmacological properties of 5-HT receptors in echinoderms is presently limited. The objective of this study was to identify the molecular and ligand activity characterisation of sea cucumber 5-HT4 receptor.
Primary identification and pharmacological characterisation of the 5-HT4 receptor was reported in 1988 through the activation of ligand 5-HT in cultured mouse embryonic collicular neurones25. Following this initial report, numerous selective 5-HT4 receptor agonists have been recognized that target 5-HT4 receptors in the central nervous system and peripheral organs and tissues26,27,28, and which thus have great therapeutic potential to treat patients suffering from a variety of diseases. In different peripheral organs, including the gastro intestinal (GI) tract, heart, and lung, 5-HT4 receptor activation is involved in various physiological processes29,30,31. For example, 5-HT4 receptor activation mediates peristaltic reflex activity due to its impaction on the enteric cholinergic axis32 and reversion of respiratory depression in etorphine-immobilisation33.
The sea cucumber Apostichopus japonicus (Selenka), is a temperate marine invertebrate animal in the family Stichopodidae whose growth is influenced by variation in water temperature34. When seawater temperature rises to a certain level during summer, most individuals of A. japonicus migrate to deeper water where they stop moving and feeding, and enter a dormant state known as aestivation. Aestivation of A. japonicus can last for as long as 4 months each year, and, in some regions of China, an aestivated A. japonicus can lose approximately 30–50% of its body mass35. During this inactive period, the rate of oxygen consumption is extremely depressed and associated with obvious hypometabolism36. Recent research, including transcriptomic, proteomic, and epigenetic, has characterised functional genes in active and aestivating A. japonicus, and has begun to reveal the mechanisms behind this process37,38,39,40,41, which has greatly advanced our understanding.
To our knowledge, little is currently known about the regulatory role of 5-HT4R in the extensive physiological adjustment during A. japonicus aestivation. In this present study, we report for the first time cloning and functional characterisation of a putative 5-HT4 receptor from the A. japonicus. Our results indicate that upon stimulation with 5-HT, the putative 5-HT4 receptor induces intracellular cAMP accumulation in dose-dependent manner and undergoes rapid internalisation mammalian cell line. Further investigation of the transcriptional expression profiles of this putative 5-HT4R between active and aestivated A. japonicus and phosphofructokinase (AjPFK) in 5-HT administrated A. japonicus in three peripheral tissues (intestine, muscle and respiratory tree) indicates that Aj5-HT4R is potentially associated with respiratory depression in aestivated A. japonicus.
Results
Isolation and Characterisation of Aj5-HT4R cDNA
The full-length 1,544 bp Aj5-HT4R cDNA sequence cloned from A. japonicus contains an open reading frame (ORF) 1,011 bp in length encoding 336 amino acids, a 5′ untranslated region (UTR) 233 bp in length, and 3′ UTR 300 bp in length (Fig. 1). The cDNA sequence was submitted to the NCBI GenBank under accession no. KX583229. One potential polyadenylation signal was identified within the 3′ UTR. The putative protein was predicted to have a molecular mass of 37.22 kDa and an isoelectric point (pI) of 9.29. The amino acid sequence of Aj5-HT4R contains the following potential sites: one typical N-glycosylation site (N87) within EC1, two conserved cysteine residues (C97 and C177) within EC1 and EC2, and 16 phosphorylation sites at 11 serine (S10, S14, S129, S131, S132, S138, S181, S186, S226, S313, and S333), four threonine (T7, T58, T229, and T332), and one tyrosine (Y133) residues (Fig. 1). The predication of 7tm_1 domain of this putative amino acid sequence indicated that Aj5-HT4R was a member of the rhodopsin-type (class A) GPCR family.

The seven transmembrane domains (TM1-TM7) are noted by the black underline. The N-glycosylated sites are highlighted in gray. The phosphorylation sites are labeled in box with full lines. The initiation codon (ATG) and the termination codon (TGA) are shown in bold. The potential polyadenylation signal (AATAAA) is noted by the double underscore. The numbers on the left refer to the position of the nucleotides and the amino acids.
The deduced amino acid sequence of A. japonicus 5-HT4R was compared with five other homologous 5-HT4R sequences, which vary in length from 351 to 388 amino acid residues. Pairwise ClustalW analysis of these amino acid sequences was carried out to evaluate homologous relationships. The predicted A. japonicus 5-HT4R amino acid sequence showed similarity to Strongylocentrotus purpuratus putative 5-HT4R sequence (40% identity), and other reported 5-HTR amino acid sequences, albeit with lower levels of similarity (from 22% to 27% identity). Multiple sequence alignment analysis revealed conservation in the 7tm_1 domain of HT4R sequences from various species including A. japonicus (Fig. 2). Apart from A. japonicus compared to S. purpuratus 5-HT4R, significant variation was observed in the length of C terminal regions (Fig. 2). Protein structure of Aj5-HT4R was predicted using SWISS-MODEL (Fig. 3A), whilst secondary structure was predicted by PredictProtein (Fig. 3B). Homology modelling revealed that this protein was similar to 4nc3.1.A in the Protein Data Bank. The protein binding region sites were predicted and marked on the constructed model.

Sequences of Strongylocentrotus purpuratus 5-HT4 receptor (Sp5-HT4R), Aplysia californica 5-HT4 receptor (Ac5-HT4R), Austrofundulus limnaeus 5-HT4 receptor (Al5-HT4R), Cavia porcellus 5-HT4 receptor (Cp5-HT4R) and Homo sapiens 5-HT4 receptor (Hs5-HT4R) were obtained from GenBank, along with a list of accession numbers (Suppl, Table S1). Alignment was generated using ClustalW and color align property was generated using Sequence Manipulation Suite online. The seven transmembrane domains (TM1–TM7) are marked with a black horizontal line above the sequence alignment. The three extracellular (EC) and three intracellular (IC) rings are noted above the sequence alignment. Black ↓ indicates the conserved cysteine residues. Percentage of sequences that must agree for identity or similarity coloring was set as 80%.

(A) Predicted 3D structure of the Aj5-HT4R protein. Seven transmembrane domains (TM1-TM7), three extracellular (EC) rings and three intracellular (IC) rings are marked. The predicted 3D structure of the Aj5-HT4R protein was generated from SWISS-MODEL. (B) Aj5-HT4R protein binding domain and transmembrane region. The Dashboard overview was generated from PredictProtein.
To examine the relationship of Aj5-HT4R with 5-HTRs from various other species, a phylogenetic tree was constructed with Mega 6.0 using ClustalW multiple alignment and protein sequences of Aj5-HT4R and 32 alternate 5-HTRs that existed in the gene bank (Fig. 4). This revealed that 5-HTRs could be divided into six groups: 5-HT1, 5-HT2, 5-HT4, 5-HT5, 5-HT6, and 5-HT7 receptors. The deduced Aj5-HT4R protein sequence was positioned alongside 5-HT4 receptors in the phylogenetic tree, which, apart from the echinoderm 5-HT4R sequence, all grouped together.

The tree was constructed based neighbor-joining algorithms using MEGA 6.0. The topological stability of the NJ tree was achieved by running 1000 bootstrapping replications. Bootstrap values (%) are indicated by numbers at the nodes. The numerical number in parentheses showed the number of sequences used in each taxon. The GenBank accession numbers and identities are listed in Suppl. Table S2.
Cellular localisation and internalisation of Aj5-HT4R
To analyse the possible sub-cellular localisation of Aj5-HT4R, the Aj5-HT4R-EGFP vector was constructed which expressed Aj5-HT4R with enhanced green fluorescent protein (EGFP) fused to the C-terminus. HEK293 cells were transiently transfected with the Aj5-HT4R-EGFP vector and GFP signal was analysed by confocal microscopy. A high level of specific fluorescence was observed in a substantial proportion of transfected cells 48 h post transfection following a 24 h starvation period in serum-free medium. As expected, fluorescence was detected on cells transfected with the Aj5-HT4R-EGFP plasmid with significant cell surface expression apparent and minimal intracellular accumulation in the absence of 5-HT (Fig. 5A).

(A) Aj5-HT4R distribution in HEK293 cells. Cells were stained with a membrane plasma probe (DiI) and a nuclei probe (DAPI). Cells stably expressing Aj5-HT4R-EGFP were seeded on glass bottom six-well plates overnight, incubated with DiI (10 μM) and DAPI, and examined by confocal microscopy as described in the section Methods. (B) Internalisation of Aj5-HT4R in HEK293 cells. HEK293 cells transfected with Aj5-HT4R-EGFP were activated by treatment with 100 pM, 10 nM and 1 μM 5-HT during 60 min and detected by confocal microscopy. The “CTL” refers to control without 5-HT stimulation. All images represent at least three independent experiments.
In order to visualise the internalisation and trafficking of Aj5-HT4R, HEK293 cells expressing Aj5-HT4R-EGFP were treated with varying concentrations of 5-HT. As described above, in the absence of 5-HT, fluorescence of Aj5-HT4R-EGFP was mainly localised in the plasma membraneand only marginally in intracellular vesicles (Fig. 5B). Following 1 μM 5-HT treatment, Aj5-HT4R-EGFP was rapidly and efficiently redistributed in the cytoplasm with distinct perinuclear accumulation (Fig. 5B), an effect that was not observed following treatment with 5-HT at lower concentrations (100 pM and 10 nM).
cAMP accumulation in Aj5-HT4R expressing cells stimulated by 5-HT
cAMP accumulation depends upon the coupling of Aj5-HT4R to G proteins. Following 1 μM 5-HT treatment of HEK293 cells transfected with pCRE-Luc vector or co-transfected with pCRE-Luc and pCMV-Flag vectors, no change in cAMP level was detected. Conversely, cAMP levels were significantly elevated following a similar treatment of HEK293 cells co-transfected with Flag-Aj5-HT4R and pCRE-Luc (Fig. 6A). To evaluate the dose-response relationship between 5-HT and Aj5-HT4R in stimulating cAMP production, HEK293 cells co-transfected with Flag-Aj5-HT4R and pCRE-Luc were treated with different 5-HT concentrations, which showed that intracellular cAMP accumulated in a dose-dependent manner with an EC50 of 53.28 ± 9.51 nM (Fig. 6B).

(A) cAMP accumulation in HEK293 cells transientlyco-transfected with Flag-Aj5-HT4R and pCRE-Luc, and was determined in response to 5-HT treatment (1 μM). (B) cAMP accumulation in HEK293 cells stably expressing Flag-Aj5-HT4R/CRE-Luc was assayed in response to different doses of 5-HT. Data are expressed as the mean ± S.E. (n = 3).
Expressional quantification of Aj5-HT4R mRNA and AjPFK mRNA
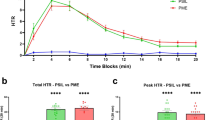
Real-time quantitative PCR assays were conducted to examine the expression patterns of Aj5-HT4R and AjPFK in various tissues of adult A. japonicus. Gene expression was normalised against expression of the house-keeping gene, β-actin (ACTB) and β-tubulin (TUBB). Aj5-HT4R mRNA was ubiquitously detected in respiratory tree, intestine, and muscle tissues of active and aestivated sea cucumbers. Aj5-HT4R expression was lower in intestine and muscle tissue during aestivation, although this difference was not statistically significant (Fig. 7A). An extremely significant decrease in Aj5-HT4R expression (p < 0.01), however, was observed in respiratory tree tissue (Fig. 7A). Moreover, to further evaluate the possible physiological role of Aj5-HT4R in the regulation of aestivation, the direct effect of 5-HT administration on the AjPFK mRNA expression was investigated. 5-HT was administrated to the aestivated A. japonicas by intraperitoneal injection, and then real-time PCR was used to quantitatively analyse the transcriptional level of AjPFK in respiratory tree, intestine, and muscle tissues. As shown in Fig. 7B, the experimental group with 5-HT administration exhibited a significant increase in AjPFK mRNA in the muscle than that of the control group with vehicle (p < 0.05).

(A) Relative expression of Aj5-HT4R in different tissues of active and aestivating sea cucumbers. Total RNA was isolated and purified from the respiratory tree (RT), intestine (IT) and muscle (MS). The expression value was normalised against the expression of the internal control gene (β-actin and β-tubulin). Each symbol and verticalbar represents mean ± SD (n = 6). Double asterisk above the bars indicates extremely significant differences (P < 0.01) between active and aestivation. (B) Tanscriptional variation of AjPFK in different tissues of 5-HT administrated sea cucumbers. Total RNA was isolated and purified from the respiratory tree (RT), intestine (IT) and muscle (MS). The expression value was normalised against the expression of the internal control gene (β-actin and β-tubulin). Each symbol and verticalbar represents mean ± SD (n = 6). Asterisk above the bars indicates extremely significant differences (P < 0.05) between control (PBS) and experimental (5-HT) groups.
Discussion
Serotonin (5-HT) produces its diverse effects through a variety of membrane-bound receptors in the central and peripheral system, and in the periphery such as the gut, cardiovascular system and blood42,43. 5-HT receptors are well-documented in vertebrates, especially in humans44. However, 5-HT receptors in marine invertebrates are comparably less characterised. In order to obtain further insight into serotonergic signaling and its functions in the echinoderm A. japonicus, we identified and functionally characterised the 5-HT4 receptor in this organism.
The putative 5-HT4R cDNA sequence cloned from A. japonicus encodes a 336 amino acid mature protein. The mature protein contains two conserved cysteine residues, a potential N-linked glycosylation site and 16 potential phosphorylation sites, which are thought to be involved in the regulation of protein trafficking and localisation, function and diversity, and signal transduction45,46. The alignment of Aj5-HT4R protein sequence and other species showed that Aj5-HT4R has the highest identity (40%) to the putative 5-HT4R of S. purpuratus. This is consistent with the evolutionary pattern of these two species. Moreover, sequence analysis revealed that the putative Aj5-HT4R together with its orthologs from other species contain amino acid motif NPxxY located at the end of the seventh transmembrane domain, which is a highly conserved among GPCRs; while unlike most of the rhodopsin-type (class A) GPCRs, the putative Aj5-HT4R and Sp5-HT4R contain a variant of the consensus D/E-R-Y/F motif with the replacement of aspartate residue with asparagines. The D/E-R-Y/F motif located between the TM3 and the second intracellular loop is a highly conserved motif in the majority of the class A GPCRs. Previous studies have indicated that both negatively charged residue D/E and positively charged residue R of the D/E-R-Y/F are involved in governing receptor conformation and G protein coupling/recognition47. The murine cytomegalovirus (MCMV) gene M33 and rat cytomegalovirus (RCMV) gene R33 encode proteins homologous to cellular chemokine receptors48, and both contain a variant NRY motif49. Studies using mutational analysis have demonstrated that the D/E residue of the D/E-R-Y/F motif is involved in agonist-dependent and independent activation of receptor response50,51,52,53. However, more efforts are required for elucidation of the function of the NRF motif of Aj5-HT4R.
Phylogenetic analysis was carried out to investigate the evolutionary relationship between 5-HTRs except for 5-HT3R which belonged to ligand-gated ion channel. The resulting phylogenetic tree revealed segmentation into six main groups (Fig. 4). Two groups comprise of receptors coupled to Gi proteins (5-HT1 and 5-HT5 receptors), three groups comprise of receptors coupled to Gs proteins (5-HT4, 5-HT6 and 5-HT7 receptors) and one group comprises of receptors coupled to Gq proteins (5-HT2 receptors) which is close to the 5-HT6 receptors group. The Aj5-HT4R clustered together with the putative 5-HT4R from S. purpuratus and was located distant to vertebrate 5-HT4Rs. Also, interestingly, the protostomian 5-HT4R from Aplysia californica, whose function in protein kinase C (PKC) activation was established by experimental data23, is also clustered in the 5-HT4R group and even closer to vertebrate 5-HT4Rs compared to echinoderm 5-HT4Rs. This suggests a common ancestor for 5-HT4Rs in deuterostomia and protostomia. Nevertheless, further experiments are required to establish the functional character and associated cell signalling pathways of echinoderm 5-HT4Rs and discover the potential difference underlying the evolutionary distance between echinoderm and vertebrate 5-HT4Rs.
In the current study, we used HEK293 cells as a heterologous expression system to functionally characterise Aj5-HT4R. We have demonstrated, for the first time, that Aj5-HT4R is activated by 5-HT with EC50 value of the response in the nanomolar range. Our data provide evidence that Aj5-HT4R is a Gs protein-coupled receptor, specifically activating adenylyl cyclase to induce intracellular cAMP formation. This is in high agreement with previous studies18,19. Additionally, to further assess Aj5-HT4R functional activity as a transmembrane receptor, fusion expression of Aj5-HT4R with the enhanced green fluorescent protein (EGFP) at the C-terminus was used to further assess Aj5-HT4R functional activity. Significant cell surface expression was observed under fluorescence microscopy (Fig. 5A), suggesting that C-terminal EGFP fusion did not affect Aj5-HT4R expression and transportation. Upon activation by agonist, fluorescence of Aj5-HT4R-EGFP was rapidly and efficiently internalised from cell surface into the cytoplasm. Agonist-induced internalisation is a well-known phenomenon for most GPCRs that control GPCR signalling, ensuring appropriate cellular responses to stimuli54. Our observation confirmed that Aj5-HT4R is a complete and functional receptor.
5-HT4 receptors are widely expressed in the body, including the central nervous system and peripheral tissues, and they exert pleiotropic effects after being activated by 5-HT. The activation of neuronal 5-HT4 receptors results in a facilitation of neurotransmitter release in the brain and the periphery. Clues to the possible functions of the 5-HT4 recptors might be obtained from its tissue distribution. In brain, 5-HT4 receptors reside in the limbic system executing their memory and learning effects55, and in hippocampus enhancing cognition and neuroprotection18. In the enteric nervous system, 5-HT4 receptor activation stimulates gastrointestinal motility in the GI tract56,57. In respiratory system, 5-HT4 receptor mediates respiratory efforts on stimulate breathing in human lung tissue31,58, and is related with pulmonary function in mice59. To investigate the potential regulatory functions of 5-HT4 receptor in A. japonicus, the expression profile of Aj5-HT4R in respiratory tree, intestine and muscle tissues of active and aestivated A. japonicus was quantitively analysed in the current study. Results indicated that the Aj5-HT4R transcript was highly expressed in respiratory tree. By contrast, we found Aj5-HT4R mRNA levels to be low in sample from all three tissues of aestivated individuals, and the lowest expression was detected in respiratory tree tissue. This is consistent with the observation of the respiratory depression of A. japonicus during aestivation36,60, it suggests that the reduction of Aj5-HT4R expression is likely related to the respiratory depression of A. japonicus during aestivation. In addition, we also demonstrated that intraperitoneal administration of 5-HT resulted in a significant increase of AjPFK expression. PFK is a rate limiting enzyme of glycolysis, and plays a key regulatory role in metabolic regulation61. It has been demonstrated that 5-HT involved in glucose uptake in skeletal muscle and regulate the glycolytic metabolism62. Although our data derived from AjPFK expression analysis do not exclude the possible roles of other 5-HT receptors in the modulation of the A. japonicas aestivation, together with the observation of Aj5-HT4R expression pattern in respiratory tree, it suggests that 5-HT/Aj5-HT4R system is likely involved in the metabolic regulation of the A. japonicas aestivation. However, more efforts are required to clarify its functional role of Aj5-HT4R in the regulation of the respiratory depression.
In conclusion, the full-length Aj5-HT4R cDNA sequence has been identified and its resulting protein has been functionally characterised in the present study. The transmembrane nature of Aj5-HT4R has been demonstrated, and internalisation of this receptor was activated by direct interaction with 5-HT. Intracellular cAMP accumulation in response to 5-HT treatment ina dose-dependent manner has been verified. The transcriptional decrease of Aj5-HT4R in respiratory treeduring aestivating periods suggests the potential relation between 5-HT4R and respiratory depression of A. japonicus during aestivation. On the other aspect, the preliminary estimated pharmacological profile of 5-HT in aestivating A. japonicus indicates the functional activity of 5-HT in metabolic regulation and suggests the potential invlovement of 5-HT receptors on respiratory control. The results presented here lead to a basic understanding of 5-HT4R in A. japonicus and further experiments should be conducted to clarify associated signalling pathways and the physiological function of this receptor.
Methods
Sample Collection
For cDNA cloning and Aj5-HT4R expression analysis in various tissues, adult individuals of the sea cucumber A. japonicus were collected separately from culture ponds in Qingdao (Shandong, China in April and June 2016). Each batch was acclimated in seawater aquaria (salinity range: 32.41–34.37). Individuals in the active group (79.3 ± 5.1 g body mass) were kept at a constant temperature (16.0 ± 0.5 °C) and fed with a formulated diet (45% marine mud, 50% Sargasso, and 5% shrimp shell powder). Individuals in the aestivating group (68.9 ± 3.2 g body mass) were also kept at a constant temperature (25.0 ± 0.5 °C) to maintain aestivation. After 15 days, respiratory tree, intestine and muscle tissues were sampled from six individuals for both groups and stored in liquid nitrogen for future use. For the experiment of adding 5-HT in aestivated sea cucumbers, intraperitoneal injection with 5 mM 5-HT (1 μL/g body weight) was carried out every 24 hours (PBS injection for control), and their respiratory tree, intestine and muscle tissues were sampled 15 days later for gene expressional quantification of phosphofructokinase (AjPFK, accession number KT779933) in glycolytic metabolism pathway.
Preparation of cDNA and Rapid amplification of cDNA ends (RACE)
Total RNA was isolated from intestine, respiration tree and muscle tissue of A. japonicus using TRIzol reagent (TaKaRa, Kusatsu, Japan) and phenol chloroform. The integrity of the total RNA was verified by electrophoresis, and RNA concentration and quality were determined using a Nanodrop 2000 (Thermo Fisher Scientific).
10 μg and 1 μg total intestine RNA was obtained to conduct 5′RLM-RACE and 3′RLM-RACE protocols, respectively. These were conducted using a FirstChoice® RLM-RACE kit (Ambion Inc., TX, USA) following manufacturer’s instructions, and samples were then stored at −20 °C for the subsequent PCR step.
For each sample, 1 μg total RNA was reverse transcribed into single-stranded cDNA by incubating with M-MLV reverse transcriptase and oligo(dT)20 (Promega Inc., Shanghai, China) at 42 °C for 1 h. An RNase inhibitor (Promega Inc., Shanghai, China) was used during cDNA synthesis to avoid RNA degradation. The cDNA was kept at −20 °C for real-time PCR. The gene-specific primers (listed in Table 1) were designed according to the partial coding sequence of Aj5-HT4R from the expressed sequence tag (EST) library, which was constructed by Trinity RNA-Seq Assembly using seven pubilshed transcriptome databases (SRR1185973, SRR1139215, SRR414930, SRR414929, SRR414928, SRR414927, SRR414926).
Sequences characterisation and phylogenetic relationships
The Aj5-HT4R cDNA sequence was used to query known sequences in GenBank using the blastx utility, BLASTX 2.2.29+ (http://blast.ncbi.nlm.nih.gov/). The full length cDNA sequence of A. japonicus 5-HT4 receptor was translated into the predicted amino acid sequence with DNAMAN 8.0. Physicochemical properties of proteins depended on Protparam (http://www.expasy.org/tools/protparam.html). N-glycosylation and phosphorylation sites were located with the NetNGlyc1.0 Server (http://www.cbs.dtu.dk/services/NetNGlyc/) and NetPhos 2.0 Server (http://www.cbs.dtu.dk/services/NetPhos/), respectively. Analysis of transmembrane in the protein was achieved by Tmpred (http://www.ch.embnet.org/software/TMPRED_form.html). Protein domains were predicted with InterProScan software (http://www.ebi.ac.uk/interpro/search/sequence-search) and SMART (http://smart.embl-heidelberg.de/). Analysis of secondary structure was predicted with PredictProtein (http://www.predictprotein.org/). The deduced amino acid sequences were aligned using ClustalW. Color Align Property was generated by Sequence Manipulation Suite (http://www.bioinformatics.org/sms2/color_align_prop.html). Aj5-HT4R protein structure was predicted using SWISS-MODEL (http://swissmodel.expasy.org/). Phylogenetic tree construction was based on the Neighbor-Joining (NJ) Method of Molecular Evolutionary Genetics Analysis (MEGA 6.0). The bootstrap value was repeated 1000 times to obtain the confidence value for the analysis.
Construction of the Mammalian Expression Vectors
To construct the Aj5-HT4R plasmid, reverse transcript PCR (RT-PCR) was performed as described in method of “Preparation of cDNA”. To amplify the CDS of Aj5-HT4R, forward primer (Aj5-HT4R-vec-F) and reverse primers (Aj5-HT4R-vec-R-EGFP) and (Aj5-HT4R-vec-R-Flag) were designed based on the full-length cDNA sequence and to allow for subcloning into the pEGFP-N1 and pCMV-Flag plasmids, respectively (Table 1). The pEGFP-N1 and pCMV-Flag vectors were purchased from Clontech Laboratories, Inc. (Palo Alto, CA), and Sigma (St.Louis, MO), respectively. The PCR products were inserted into the final pEGFP-N1 and pCMV-Flag expression vectors using the EcoRI and KpnI restriction enzymes (Beyotime, Haimen, China) and Rapid DNA Ligation Kit (Beyotime, Haimen, China). The constructed vectors were named Aj5-HT4R-EGFP and Flag-Aj5-HT4R, respectively, and sequenced to verify sequence fidelity, orientation, and reading frame.
Transfection
The human embryonic kidney cell line (HEK293) was maintained in Dulbecco’s modified Eagle’s medium (DMEM, Hyclone, Logan, UT, USA) supplemented with 10% foetal bovine serum (FBS, HyClone, Logan, UT, USA) and 4 mM L-glutamine (Invitrogen, Madison, WI, USA). The Aj5-HT4R-EGFP and Flag-Aj5-HT4R vectors were transfected into HEK293 cells using Lipofectamine 2000 (Invitrogen, Madison, WI, USA) according to manufacturer’s instructions.
Internalisation assay and Confocal Microscopy
HEK293 cells expressing Aj5-HT4R-EGFP were seeded onto glass coverslips coated with 0.1 mg/ml of poly-L-lysine and allowed to attach overnight under normal growth conditions. After 24 h, the HEK293 cells starved for a further 24 h in serum-free medium to eliminate the effects of endogenous 5-HT in FBS. For receptor surface expression analysis, cells were stained with the membrane probe DiI (Beyotime, Haimen, China) at 37 °C for 5–10 min, fixed with 4% paraformaldehyde for 15 min, and finally incubated with DAPI (Beyotime, Haimen, China) for 10 min. For the internalisation assay, cells expressing Aj5-HT4R-EGFP were treated with 100 pM, 10 nM and 1 μM 5-HT (sigma, Saint Louis, USA) for 60 min at 37 °C, then fixed with 4% paraformaldehyde for 15 min. Cells were visualized by fluorescence microscopy on a Leica TCS SP5II laser scanning confocal microscope using a HCX PL APO lambda blue 63× 1.4 oil immersion lens.
cAMP accumulation measurement
After seeding in a 96-well plate overnight, HEK293 cells stably cotransfected with Flag-Aj5-HT4R and pCRE-Luc vectors were grown to 80–85% confluence, before being starved for 24 h in serum-free medium to eliminate the effects of the endogenous 5-HT in FBS. Cells were thentreated with the indicated concentration of 5-HT(1 pM, 10 pM, 100 pM, 1 nM, 10 nM, 100 nM, 1 mM, and 10 mM) in DMEM without FBS and incubated for 4 h at 37 °C, with application repeated a total of three times. Luciferase activity was detected using a firefly luciferase assay kit (Kenreal, Shanghai, China).
Real-time quantitative PCR (qRT-PCR)
For qRT-PCR, β-actin (ACTB) and β-tubulin (TUBB) were chosen as the internal control (housekeeping) genes and gene-specific primers were designed based on the ORF sequences63,64. Specific qRT-PCR primers for Aj5-HT4R and AjPFK were designed based on CDS (Table 1). The primers were tested to ensure amplification of single discrete bands with no primer-dimers. qRT-PCR assays were carried out using the SYBR PrimeScript™ RT reagent Kit (TaKaRa, Kusatsu, Japan) following manufacturer’s instructions, and ABI 7500 Software v2.0.6 (Applied Biosystems, UK). qRT-PCR was performed for 35 cycles with the following condition: 95 °C/5 s, 60 °C/30 s. The relative level of gene expression was calculated using the 2−△△Ct method and data was normalised by geometric averaging of the internal control genes65,66. Differences between experimental and control groups were tested using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test, using PASW Statistics 18.00 (SPSS Inc., Chicago, IL, USA). Significance was set at P < 0.05, and extremely significance was set at P < 0.01.
Additional Information
How to cite this article: Wang, T. et al. Identification and functional characterisation of 5-HT4 receptor in sea cucumber Apostichopus japonicus (Selenka). Sci. Rep. 7, 40247; doi: 10.1038/srep40247 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Rapport, M. M., Green, A. A. & Page, I. H. Serum vasoconstrictor, serotonin; isolation and characterization. J Biol Chem 176, 1243–1251 (1948).
Barnes, N. M. 5-HT: the promiscuous and happy hormone! Curr Opin Pharmacol 11, 1–2 (2011).
Hayes, D. J. & Greenshaw, A. J. 5-HT receptors and reward-related behaviour: a review. Neurosci Biobehav Rev 35, 1419–1449 (2011).
Berger, M., Gray, J. A. & Roth, B. L. The expanded biology of serotonin. Annual review of medicine 60, 355–366 (2009).
Yao, K. et al. Tryptophan metabolism in animals: important roles in nutrition and health. Front Biosci (Schol Ed) 3, 286–297 (2011).
Wirth, A., Holst, K. & Ponimaskin, E. How serotonin receptors regulate morphogenic signalling in neurons. Progneurobiol (2016).
Collingridge, G. L., Olsen, R. W., Peters, J. & Spedding, M. A nomenclature for ligand-gated ion channels. Neuropharmacology 56, 2–5 (2009).
Hoyer, D., Hannon, J. P. & Martin, G. R. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol biochem be 71, 533–554 (2002).
Hannon, J. & Hoyer, D. Molecular biology of 5-HT receptors. Behav Brain Res 195, 198–213 (2008).
Blondel, O., Gastineau, M., Dahmoune, Y., Langlois, M. & Fischmeister, R. Cloning, expression, and pharmacology of four human 5-hydroxytryptamine 4 receptor isoforms produced by alternative splicing in the carboxyl terminus. J Neurochem 70, 2252–2261 (1998).
Zhu, T. et al. Expression of serotonin receptors in the colonic tissue of chronic diarrhea rats. Saudi J Gastroenterol 22, 234–239 (2016).
dos Santos, T. S. et al. Distribution of serotonin 5-HT1A-binding sites in the brainstem and the hypothalamus, and their roles in 5-HT-induced sleep and ingestive behaviors in rock pigeons (Columba livia). Behav Brain Res 295, 45–63 (2015).
Perez-Maceira, J. J., Otero-Rodino, C., Mancebo, M. J., Soengas, J. L. & Aldegunde, M. Food intake inhibition in rainbow trout induced by activation of serotonin 5-HT2C receptors is associated with increases in POMC, CART and CRF mRNA abundance in hypothalamus. J Comp Physiol B 186, 313–321 (2016).
Komuniecki, R. W., Hobson, R. J., Rex, E. B., Hapiak, V. M. & Komuniecki, P. R. Biogenic amine receptors in parasitic nematodes: what can be learned from Caenorhabditis elegans? Mol Biochem Parasitol 137, 1–11 (2004).
Spitzer, N., Edwards, D. H. & Baro, D. J. Conservation of structure, signaling and pharmacology between two serotonin receptor subtypes from decapod crustaceans, Panulirus interruptus and Procambarus clarkii . J Exp Biol 211, 92–105 (2008).
Blenau, W. & Thamm, M. Distribution of serotonin (5-HT) and its receptors in the insect brain with focus on the mushroom bodies: lessons from Drosophila melanogaster and Apis mellifera . Arthropod Struct Dev 40, 381–394 (2011).
Thamm, M. et al. Function and distribution of 5-HT2 receptors in the honeybee (Apis mellifera). Plos One 8, e82407 (2013).
Berumen, L. C., Rodriguez, A., Miledi, R. & Garcia-Alcocer, G. Serotonin receptors in hippocampus. TheScientificWorldJournal 2012, 823493 (2012).
Vleugels, R., Verlinden, H. & Broeck, J. V. Serotonin, serotonin receptors and their actions in insects. Neurotransmitter 2, 1–14 (2015).
Qi, Y. X. et al. Larvae of the small white butterfly, Pieris rapae, express a novel serotonin receptor. J Neurochem 131, 767–777 (2014).
Kawahara, H., Isoai, A. & Shizuri, Y. Molecular cloning of a putative serotonin receptor gene from barnacle, Balanus amphitrite. Gene 184, 245–250 (1997).
Lee, Y. S. et al. Identification of a serotonin receptor coupled to adenylyl cyclase involved in learning-related heterosynaptic facilitation in Aplysia . Proc Natl Acad Sci USA 106, 14634–14639 (2009).
Nagakura, I. et al. Regulation of protein kinase C Apl II by serotonin receptors in Aplysia. J Neurochem 115, 994–1006 (2010).
Katow, H., Yaguchi, S., Kiyomoto, M. & Washio, M. The 5-HT receptor cell is a new member of secondary mesenchyme cell descendants and forms a major blastocoelar network in sea urchin larvae. Mech Dev 121, 325–337 (2004).
Dumuis, A., Bouhelal, R., Sebben, M., Cory, R. & Bockaert, J. A nonclassical 5-hydroxytryptamine receptor positively coupled with adenylate cyclase in the central nervous system. Mol Pharmacol 34, 880–887 (1988).
De Maeyer, J. H., Lefebvre, R. A. & Schuurkes, J. A. 5-HT4 receptor agonists: similar but not the same. Neurogastroenterol Motil 20, 99–112 (2008).
Manabe, N., Wong, B. S. & Camilleri, M. New-generation 5-HT4 receptor agonists: potential for treatment of gastrointestinal motility disorders. Expert Opin Investig Drugs 19, 765–775 (2010).
Nirogi, R. et al. Synthesis and SAR of Imidazo[1,5-a]pyridine derivatives as 5-HT4 receptor partial agonists for the treatment of cognitive disorders associated with Alzheimer’s disease. European journal of medicinal chemistry 103, 289–301 (2015).
Priem, E., Van Colen, I., De Maeyer, J. H. & Lefebvre, R. A. The facilitating effect of prucalopride on cholinergic neurotransmission in pig gastric circular muscle is regulated by phosphodiesterase 4. Neuropharmacology 62, 2126–2135 (2012).
De Maeyer, J. H., Schuurkes, J. A. & Lefebvre, R. A. Selective desensitization of the 5-HT4 receptor-mediated response in pig atrium but not in stomach. Br J Pharmacol 156, 362–376 (2009).
Richter, D. W., Manzke, T., Wilken, B. & Ponimaskin, E. Serotonin receptors: guardians of stable breathing. Trends Mol Med 9, 542–548 (2003).
Gershon, M. D. & Tack, J. The serotonin signaling system: from basic understanding to drug development for functional GI disorders. Gastroenterology 132, 397–414 (2007).
Meyer, L. C., Hetem, R. S., Fick, L. G., Mitchell, D. & Fuller, A. Effects of serotonin agonists and doxapram on respiratory depression and hypoxemia in etorphine-immobilized impala (Aepyceros melampus). J Wildl Dis 46, 514–524 (2010).
Zhang, L., Pan, Y. & Song, H. Environmental Drivers of Behavior. Developments in Aquaculture and Fisheries Science 39, 133–152 (2015).
Yang, H. S. et al. Effects of body size and water temperature on food consumption and growth in the sea cucumber Apostichopus japonicus (Selenka) with special reference to aestivation. Aquac Res 36, 1085–1092 (2005).
Yang, H. S. et al. Metabolic characteristics of sea cucumber Apostichopus japonicus (Selenka) during aestivation. J Exp Mar Bio Ecol 330, 505–510 (2006).
Du, H. et al. Transcriptome sequencing and characterization for the sea cucumber Apostichopus japonicus (Selenka, 1867). Plos One 7, e33311 (2012).
Chen, M. Y., Zhu, A. & Storey, K. B. Comparative phosphoproteomic analysis of intestinal phosphorylated proteins in active versus aestivating sea cucumbers. J Proteomics (2015).
Zhao, Y., Yang, H. S., Storey, K. B. & Chen, M. Y. RNA-seq dependent transcriptional analysis unveils gene expression profile in the intestine of sea cucumber Apostichopus japonicus during aestivation. Comp Biochem Physiol Part D Genomics Proteomics 10, 30–43 (2014).
Zhao, Y., Chen, M. Y., Storey, K. B., Sun, L. & Yang, H. S. DNA methylation levels analysis in four tissues of sea cucumber Apostichopus japonicus based on fluorescence-labeled methylation-sensitive amplified polymorphism (F-MSAP) during aestivation. Comp Biochem Physiol B Biochem Mol Biol 181, 26–32 (2015).
Zhao, P. et al. Identification of differential expressed proteins and characterization their mRNA expression in thermally stressed Apostichopus japonicus . Comp Biochem Physiol Part D Genomics Proteomics 8, 194–200 (2013).
Liu, H. N. et al. Serotonin augments gut pacemaker activity via 5-HT3 receptors. Plos One 6, e24928 (2011).
Kaumann, A. J. & Levy, F. O. 5-hydroxytryptamine receptors in the human cardiovascular system. Pharmacol Ther 111, 674–706 (2006).
Pytliak, M., Vargova, V., Mechirova, V. & Felsoci, M. Serotonin receptors - from molecular biology to clinical applications. Physiol Res 60, 15–25 (2011).
Bray, L. et al. Identification and functional characterization of the phosphorylation sites of the neuropeptide FF2 receptor. J Biol Chem 289, 33754–33766 (2014).
Arnold, J. N., Wormald, M. R., Sim, R. B., Rudd, P. M. & Dwek, R. A. The impact of glycosylation on the biological function and structure of human immunoglobulins. Annu Rev Immunol 25, 21–50 (2007).
Rovati, G. E., Capra, V. & Neubig, R. R. The highly conserved DRY motif of class A G protein-coupled receptors: beyond the ground state. Mol Pharmacol 71, 959–964 (2007).
Chee, M. S., Satchwell, S. C., Preddie, E., Weston, K. M. & Barrell, B. G. Human cytomegalovirus encodes three G protein-coupled receptor homologues. Nature 344, 774–777 (1990).
Case, R. et al. Functional analysis of the murine cytomegalovirus chemokine receptor homologue M33: ablation of constitutive signaling is associated with an attenuated phenotype in vivo . J Virol 82, 1884–1898 (2008).
Scheer, A., Fanelli, F., Costa, T., De Benedetti, P. G. & Cotecchia, S. The activation process of the alpha1B-adrenergic receptor: potential role of protonation and hydrophobicity of a highly conserved aspartate. Proc Natl Acad Sci USA 94, 808–813 (1997).
Morin, D. et al. The D136A mutation of the V2 vasopressin receptor induces a constitutive activity which permits discrimination between antagonists with partial agonist and inverse agonist activities. FEBS Lett 441, 470–475 (1998).
Arora, K. K., Cheng, Z. & Catt, K. J. Mutations of the conserved DRS motif in the second intracellular loop of the gonadotropin-releasing hormone receptor affect expression, activation, and internalization. Mol Endocrinol 11, 1203–1212 (1997).
Lu, Z. L., Curtis, C. A., Jones, P. G., Pavia, J. & Hulme, E. C. The role of the aspartate-arginine-tyrosine triad in the m1 muscarinic receptor: mutations of aspartate 122 and tyrosine 124 decrease receptor expression but do not abolish signaling. Mol Pharmacol 51, 234–241 (1997).
Yang, J. W. et al. Agonist-Activated Bombyx Corazonin Receptor Is Internalized via an Arrestin-Dependent and Clathrin-Independent Pathway. Biochemistry 55, 3874–3887 (2016).
Langlois, M. & Fischmeister, R. 5-HT4 receptor ligands: applications and new prospects. J Med Chem 46, 319–344 (2003).
Bielefeldt, K., Tuteja, A. & Nusrat, S. Disorders of gastrointestinal hypomotility. F1000Res 5 (2016).
Lefebvre, R. A., Van Colen, I., Pauwelyn, V. & De Maeyer, J. H. Synergistic effect between 5-HT4 receptor agonist and phosphodiesterase 4-inhibitor in releasing acetylcholine in pig gastric circular muscle in vitro . Eur J Pharmacol 781, 76–82 (2016).
Teran, F. A., Massey, C. A. & Richerson, G. B. Serotonin neurons and central respiratory chemoreception: where are we now? Prog Brain Res 209, 207–233 (2014).
Dupont, L. L. et al. Investigation of 5-HT4 receptors in bronchial hyperresponsiveness in cigarette smoke-exposed mice. Pulm Pharmacol Ther 28, 60–67 (2014).
Zhao, Y., Yang, H. S., Storey, K. B. & Chen, M. Y. Differential gene expression in the respiratory tree of the sea cucumber Apostichopus japonicus during aestivation. Mar Genomics 18 Pt B, 173–183 (2014).
Yi, W. et al. Phosphofructokinase 1 glycosylation regulates cell growth and metabolism. Science 337, 975–980 (2012).
Coelho, W. S. & Sola-Penna, M. Serotonin regulates 6-phosphofructo-1-kinase activity in a PLC-PKC-CaMK II- and Janus kinase-dependent signaling pathway. Mol Cell Biochem 372, 211–220 (2013).
Zhu, A. J., Chen, M. Y., Zhang, X. M. & Storey, K. B. Gene structure, expression, and DNA methylation characteristics of sea cucumber cyclin B gene during aestivation. Gene (2016).
Xiang, X. W. et al. Glycolytic regulation in aestivation of the sea cucumber Apostichopus japonicus: evidence from metabolite quantification and rate-limiting enzyme analyses. Marine Biology 163, 1–12 (2016).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 25, 402–408 (2001).
Vandesompele, J. et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biology 3 (2002).
Acknowledgements
The authors of this paper would like to thank Prof. Jiayan Xie for his technical assistance and equipment usage. This work was financially supported by National Natural Science Foundation of China (41406137), Natural Science Foundation of Zhejiang Province (LY14D060002).
Author information
Authors and Affiliations
Contributions
T.W., N.Z. and C.W. conceived and coordinated the study. Z.Y. and T.W. wrote the main manuscript text, Z.Y. prepared Figures 1–4, Table 1 and supplyementary information, T.W. prepared Figures 5–7. Z.Y., T.W. and L.S. designed, performed the experiments. Z.Y., T.W., L.S. and Z.L. analysed the results. N.Z. provided technical assistance. All authors reviewed the results and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wang, T., Yang, Z., Zhou, N. et al. Identification and functional characterisation of 5-HT4 receptor in sea cucumber Apostichopus japonicus (Selenka). Sci Rep 7, 40247 (2017). https://doi.org/10.1038/srep40247
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep40247
This article is cited by
-
Effects of increasing temperature and aestivation on biogenic amines, signal transduction pathways and metabolic enzyme activities in the sea cucumber (Apostichopus japonicus)
Marine Biology (2022)
-
Fine-mapping and association analysis of candidate genes for papilla number in sea cucumber, Apostichopus japonicus
Marine Life Science & Technology (2022)
-
Distribution and dynamic expression of serotonin and dopamine in the nervous system and ovary of Holothuria scabra during ovarian maturation
Journal of Comparative Physiology A (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
