
Abstract
Receipt of broad-spectrum antibiotics enhances Candida albicans colonization of the GI tract, a risk factor for haematogenously-disseminated candidiasis. To understand how antibiotics influence C. albicans colonization, we treated mice orally with vancomycin or a combination of penicillin, streptomycin and gentamicin (PSG) and then inoculated them with C. albicans by gavage. Only PSG treatment resulted in sustained, high-level GI colonization with C. albicans. Furthermore, PSG reduced bacterial diversity in the colon much more than vancomycin. Both antibiotic regimens significantly reduced IL-17A, IL-21, IL-22 and IFN-γ mRNA levels in the terminal ileum but had limited effect on the GI fungal microbiome. Through a series of models that employed Bayesian model averaging, we investigated the associations between antibiotic treatment, GI microbiota and host immune response and their collective impact on C. albicans colonization. Our analysis revealed that bacterial genera were typically associated with either C. albicans colonization or altered cytokine expression but not with both. The only exception was Veillonella, which was associated with both increased C. albicans colonization and reduced IL-21 expression. Overall, antibiotic-induced changes in the bacterial microbiome were much more consistent determinants of C. albicans colonization than either the GI fungal microbiota or the GI immune response.
Similar content being viewed by others

Introduction
The fungus Candida albicans is a human commensal that grows on the skin and mucosal surfaces of healthy individuals1. However, in susceptible patients, C. albicans can enter the bloodstream, either by translocation across the mucosa of the gastrointestinal (GI) tract2 or via an indwelling vascular catheter3. The resulting haematogenously-disseminated infection is associated with a mortality of up to 49% and is the fourth most common cause of hospital-acquired bloodstream infections in the U.S.3.
Risk factors for developing disseminated and invasive candidiasis include the receipt of broad-spectrum antibiotics, immunosuppression and colonization with Candida species. It is believed that broad-spectrum antibiotics predispose patients to developing disseminated candidiasis by suppressing the competing bacterial microbiota in the GI tract and encouraging the overgrowth of C. albicans2,4,5,6,7. In recent randomized clinical trials, probiotics were effective in reducing GI colonization by Candida species and decreasing the incidence of invasive candidiasis8,9. In immunodeficient and malnourished mice, introduction of Lactobacillus species into the GI tract decreased GI C. albicans burden and resulted in lower mortality10,11. These results suggest that the composition of the GI tract microbiota influences the level of C. albicans colonization.
Another potential influence on the level of C. albicans GI colonization is the host immune status12. Mutations in genes that code for host cytokines such as interleukin (IL) -17, IL-22 and interferon (IFN) -γ increase susceptibility to mucosal candidiasis in both mice and humans12,13. T-cell effector populations that drive cytokine responses express receptors for peptide antigens of specific commensal bacteria14. Among these, gram-positive bacilli from the genus Clostridium15 and the closely related genus, “Candidatus Arthromitus” also known as the segmented filamentous bacteria (SFB), are prominent activators of adaptive Th17 immunity in the GI tract16,17,18. By depleting these immune-activating bacteria2 and through possible direct immunomodulation19, antibiotics have the potential to enhance C. albicans GI colonization.
A mechanistic understanding of the tri-faceted interface of antibiotic action on the local immune response, GI bacterial and fungal microbiota and C. albicans colonization is essential for developing strategies to reduce the incidence of candidiasis. Antibiotic regimens that include penicillin are associated with substantially higher rates of GI colonization with Candida species when compared with those without penicillin20,21. Therefore, to examine the factors that influence the level of C. albicans GI colonization, we contrasted the effects of a combination of penicillin, streptomycin and gentamicin (PSG) with those of vancomycin in a mouse model. We studied two GI sites: a) the terminal ileum because the small intestine contains a large number of immune cells, particularly those involved in a Th17 response16 and b) the colon (sampled via faecal pellets), which is the most commonly sampled site for profiling the microbiome22,23. Unlike humans, mice are not normally colonized with C. albicans unless the resident GI flora is perturbed by the administration of oral antibiotics21. Hence, the mouse model provided us a controlled platform to perform our experiments.
Mouse studies that incorporate next generation sequencing (NGS) for measuring microbiome variables represent a high-dimensional setting where the number of measurements substantially exceeds the number of samples. As an example, an experiment with 30 mice can generate over 300 bacterial, fungal and immune measurements per mouse. As a consequence of this high-dimensionality, a large number of equally likely models with various combinations of microbes and other covariables can explain antibiotic effects and subsequent colonization24. However, most current microbiome studies do not systematically explore this large model space. Instead, a number of these studies apply a univariate method such as the one-way ANOVA test, Wilcoxon rank-sum test or Kruskal-Wallis rank-sum test to individually screen each measured microbiome variable for a significant effect. This significance testing typically includes correction strategies for multiple testing. For any response of interest, the effects of the significant microbes selected by the univariate method are then estimated using a single regression model25.
While univariate microbiome variable selection followed by a single regression model has yielded important new information about factors that affect the composition of microbiomes, the p-values or confidence intervals (CI) that are computed after univariate screening often overestimate the strength of conclusions26,27. Even with corrections for multiple testing in place, independent tests do not consider the complex multivariable, correlational structure of microbiome data. Furthermore, the final step of inference is based upon a single model that ignores the uncertainty that compounds with each variable selection step prior to building the final model28. Ensemble multivariable regression modelling approaches such as Bayesian model averaging (BMA) have been used in ecology to address these shortcomings29. Ensemble methods examine a large number of variable configurations to identify significant variables and to simultaneously compute their effect sizes. After a systematic evaluation of several such ensemble approaches on our mouse microbiome data, we selected BMA for its strong performance across several variable selection and ranking metrics30. In addition, BMA helped us formally account for model uncertainty in high-dimensions by simultaneously computing two crucial statistical metrics for each finding: a) consistency as measured by posterior inclusion probability (PIP) and b) statistical significance using the 95% CI of effect size31. Thus, BMA has a distinct advantage over most statistical approaches, which typically provide estimates of statistical significance but not of consistency.
We employed BMA to identify the microbiome community and the host immune response signatures that characterize antibiotic treatment and to probabilistically rank the influence of the microbiome and the host immune response on the level of C. albicans colonization of the mouse GI tract following gastric challenge with this organism. Our analysis revealed that antibiotic-induced changes in the bacterial microbiome were the most consistent determinants of C. albicans colonization. These bacterial genera typically influenced the growth of C. albicans without simultaneously altering the host GI immune response. Although antibiotics significantly altered the composition of GI fungal microbiota and suppressed the GI immune response, these factors were less influential on colonization relative to the bacterial microbiota.
Results
PSG and vancomycin had differential effects on C. albicans colonization and microbial diversity, but induced similar host immune responses
We used a mouse model to investigate the tri-faceted effect of antibiotics on the host immune response, the GI bacterial and fungal microbiota and C. albicans colonization in the GI tract. Mice in the control group received sterile drinking water, while those in treatment groups received sterile drinking water containing either vancomycin or PSG. After 7 days, we inoculated some of the mice in the control and treatment groups with 105 C. albicans cells, administered by gavage (Figure 1). We then continued the mice on their original control or treatment regimens for an additional 14 days. Each group had 3 to 7 mice and the results of at least two independent experiments were combined (details in Methods).

Experimental Design.
Following initiation of antibiotics and inoculation with C. albicans, we collected samples of the mouse terminal ileum and faecal pellets. We then sequenced the bacterial 16S and fungal ITS amplicons in the samples by 454 sequencing and taxonomically classified these sequences using a tailored bioinformatics workflow. We quantified mRNA expression levels of 6 host cytokines (C mRNA) including IL-17A, IL-21, IL-22, IFN-γ, TNF-α and IL-4 from segments of the terminal ileum. Using quantitative culture, we determined the level of C. albicans colonization in the faecal pellets in terms of colony forming units per gram (CFU/g).
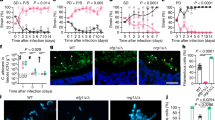
Following exposure to C. albicans, the faecal pellets of control mice contained no detectable C. albicans cells by quantitative culture on days 9, 14 and 21 (Figure 2a). In contrast, mice that received PSG had sustained, high-level of colonization with C. albicans that persisted throughout the duration of the experiment. Unlike PSG treatment, vancomycin treatment induced only a low-level, transient colonization on day 9 and no C. albicans cells were cultured from the faecal pellets on days 14 and 21.

Exploring the effects of PSG (P) and vancomycin (Van., V) relative to controls on:
(a) C. albicans colonization: Levels of log transformed colony forming units per gram (CFU/g) of C. albicans from faecal pellets sampled on days 9, 14 and 21 from controls (green), vancomycin-treated (violet) and PSG-treated (orange) mice. (b) Cytokine expression levels: mRNA expression levels of the 6 cytokines measured from segments of the terminal ileum sampled on days 7 and 21. +CA and No CA respectively indicate mice groups with and without exposure to C. albicans gavage. (c) Microbiome diversity: bacterial and fungal diversity on days 7 and 21 both with (+CA) and without C. albicans exposure (No CA) in the faecal pellets and the terminal ileum. Diversity was computed using the inverse Simpson index. Statistical analysis: In each boxplot, the upper and lower hinges correspond to the 25th and 75th percentiles of the data, also known as the interquartile range (IQR). The upper and lower whiskers extend from the hinge to the highest value within 1.5 times the IQR. Points beyond the end of the whiskers denote outliers. Asterisks, when present, denote statistically significant differences relative to controls. Findings were considered statistically significant when the 99.6% bootstrap confidence intervals (CI) of the normalized mean difference (Welch's statistic) between the antibiotic group and the controls did not include zero. The 99.6% bootstrap CIs were computed over 10,000 resamples and included a conservative correction for 6 simultaneous hypotheses corresponding to the 6 cytokines. Results shown are the combined data from at least two independent experiments of 3–5 mice each.
Next, we determined the local GI immune response in the various treatment groups using real-time PCR to measure the mRNA expression levels of six cytokines in the terminal ileum. These cytokines consisted of IL-4, IL-17A, IL-21, IL-22, IFN-γ and tumour necrosis factor (TNF) -α. We selected these cytokines because they have been shown to be relevant in the context of C. albicans colonization of the GI tract32. Relative to the controls, treatment with either vancomycin or PSG was associated with significantly depressed expression levels of IL-17A, IL-22 and IFN-γ on both days 7 and 21, both in the presence and absence of C. albicans exposure (Figure 2b).
Finally, we compared the bacterial and fungal diversity, using the inverse Simpson diversity index, in PSG and vancomycin-treated mice relative to controls. The GI bacterial and fungal microbiomes were profiled by sequencing the bacterial 16S rRNA and fungal ITS regions followed by taxonomic classification. Across our experiments, we detected 344 bacterial genera and 109 fungal genera. In the faecal pellets, both vancomycin and PSG treatments significantly decreased bacterial diversity on day 7 and day 21 in the presence of C. albicans, relative to controls. PSG treatment caused a greater reduction in diversity, as expected from the broader spectrum of activity of this antibiotic combination (Figure 2c). In the terminal ileum, treatment with vancomycin did not affect bacterial diversity, while treatment with PSG resulted in an increase in diversity, more so in the absence of C. albicans exposure (Figure 2c).
Compared to bacterial diversity, fungal diversity was lower in both the colon and terminal ileum. Two weeks after C. albicans challenge, there was an increase in fungal diversity in the vancomycin-treated mice in the faecal pellets. In contrast, the fungal diversity remained low in the PSG-treated mice (Figure 2c). In comparison to the faecal pellets, the fungal diversity in the terminal ileum of the controls was lower. Collectively, these results indicate that vancomycin and PSG had different effects on microbial diversity in the GI tract. Moreover, the effect of the antibiotics differed across locations in the GI tract.
Thus our exploratory analysis of the three facets of the microbiome-immune-colonization interface revealed that both vancomycin and PSG reduced bacterial diversity in the faecal pellets and suppressed cytokine mRNA expression in the terminal ileum. However, only PSG induced susceptibility to sustained C. albicans colonization.
The antibiotic-microbiome interface: PSG and vancomycin resulted in markedly different bacterial and fungal microbiome signatures
We next identified the set of specific bacterial and fungal genera that were significantly and consistently altered by either PSG or vancomycin relative to controls by building multivariable BMA logistic regression models (Figure 3)33. BMA explored a set of 10,000 genera configurations that differentiated PSG and vancomycin from controls. The posterior inclusion probability (PIP) for any genus under BMA is the relative frequency with which it was selected as influential across these 10,000 configurations31. Genera with higher PIPs were able to consistently differentiate antibiotics versus control status across samples.

Effect of antibiotics on the bacterial microbiome.
Results from BMA logistic regression ensembles exploring the antibiotic and bacterial microbiome interface are shown in panels (a) and (c) for the faecal pellets and (b) and (d) for the terminal ileum. Panels (a) and (b) plot the median effect size of each bacterium against its posterior inclusion probability (PIP) in the model. PIP is expressed in %. The effect size of a microbe is its regression coefficient in the model and depends on its relative abundance under antibiotic treatment (PSG or Van.) relative to controls (Cntrl), after adjusting for all the other covariables in the model. We show only the effects that were statistically significant i.e. the corresponding Bayesian 95% CIs did not include zero. Higher PIPs indicate higher consistency in antibiotic effects. PIPs can range from 0% (not consistent) to 100% (very highly consistent). All models were built at the genus level. Each point in the graph denotes a genus coloured by its phylum membership. Panels (c) and (d) show heatmaps of relative abundances (scaled to the range 0–1) of bacteria with the top consistent and significant differences across the PSG (P), vancomycin (Van., V) and control (C) groups on days 7 and 21. Each genus is annotated with its phylum-level label in the heatmaps. Numbers in square brackets denote distinct sequence clusters that were mapped to the same genus but could not be collapsed since they were more than 3% dissimilar from each other.
The effect size for a given genus is its regression coefficient in the model and is determined by the difference in its relative abundance between antibiotic treated mice and control mice, after adjusting for all the other microbe proportions and cytokine variables in the model. We built separate models to examine the effects of antibiotics on the bacterial and fungal microbiota in the faecal pellets and in the terminal ileum for days 7 and 21 (Figure 3). We show only the genera whose 95% CI of effect sizes did not include zero and thus, represent statistically significant antibiotic effects.
Overall, the maximum PIPs in the antibiotic-microbiome models were approximately 30%, indicating that neither PSG nor vancomycin had a consistent impact on any given genus across samples (Figures 3a and b). In the faecal pellets, the effects of PSG on the bacterial microbiome were the most heterogeneous, with a large number of high and significant effect sizes accompanied by relatively low PIPs (Figure 3a). Unlike PSG, vancomycin was associated with a lower number of significant effects. However, these effects were more consistent. In the terminal ileum, PSG and vancomycin had a similar pattern of effects on the bacterial microbiome and the signatures in the ileum were more consistent than those in the faecal pellets (Figure 3b). In both GI sites, bacterial genera belonging to the Bacteroidetes and Firmicutes phyla had the most consistent antibiotic signatures with higher PIPs.
In the faecal pellets, PSG consistently and significantly suppressed most genera on both days 7 and 21 (Figure 3c). Most of these genera were members of phyla Bacteroidetes and Firmicutes. By contrast, Parabacteroides was the only genus that increased in relative abundance in the PSG-treated mice on days 7 and 21. Even though vancomycin reduced the relative abundance of several of the same genera as did PSG, the magnitude of the reduction was smaller in comparison. In addition, vancomycin increased multiple genera, including Lactobacillus, Proteus, Cronobacter, Anaeroplasma, Parasutterella and Mucispirillum.
In the terminal ileum, PSG treatment had a mixed effect, decreasing the relative abundance of some genera, but increasing others (Figure 3d). The increase in Enterococcus, Streptophyta and Anaeroplasma was especially notable at day 21 in the presence of C. albicans colonization. Vancomycin also had a similar mixed effect. It depleted the same genera as did PSG, including “Candidatus Arthromitus” which is known to stimulate a Th17 response in the GI tract15,16. However, vancomycin treatment increased a greater number of genera than did PSG, especially on day 7.
On day 7, the fungal microbiome of mice receiving either PSG or vancomycin had substantial mouse-to-mouse variability in both GI sites (Figure 4a and 4b). However, by day 21, PSG induced a larger number of consistent effects in the fungal genera of the faecal pellets than did vancomycin. Five of these effects were on sequence clusters that mapped to Candida (Figure 4c). Of these, Candida [2] decreased under both antibiotics while Candida [1], [4] and [5] increased only under PSG. Candida [6] was present in a higher proportion in controls, increased under vancomycin and decreased under PSG.

Effect of antibiotics on the fungal microbiome.
Results from BMA logistic regression ensembles exploring the antibiotic and fungal microbiome interface are shown in panels (a) and (c) for the faecal pellets and (b) and (d) for the terminal ileum. Panels (a) and (b) plot the median effect size of each fungus against its posterior inclusion probability (PIP) in the model. PIP is expressed in %. The effect size of a microbe is its regression coefficient in the model and depends on its relative abundance under antibiotic treatment (PSG or Van.) relative to controls (Cntrl), after adjusting for all the other covariables in the model. We show only the effects that were statistically significant i.e. the corresponding Bayesian 95% CIs did not include zero. Higher PIPs indicate higher consistency in antibiotic effects. PIPs can range from 0% (not consistent) to 100% (very highly consistent). All models were built at the genus level. Each point in the graph denotes a genus coloured by its phylum membership. Panels (c) and (d) show heatmaps of abundances (scaled to the range 0–1) of fungi with the top consistent and significant differences across the PSG (P), vancomycin (Van., V) and control (C) groups on days 7 and 21. Each genus is annotated with its phylum-level label in the heatmaps. Numbers in square brackets denote distinct sequence clusters that were mapped to the same genus but could not be collapsed since they were more than 3% dissimilar from each other. UC denotes sequence clusters that did not have a genus level classification in any of the databases (NCBI, SILVA) we used as reference for taxonomic classification and could not be resolved using the Megablast algorithm.
In the terminal ileum, neither antibiotic had consistent effects on the fungal microbiome.
We were specifically interested in obtaining insights into Candida colonization and therefore explored the species level composition of Candida clusters that were identified as influential by the BMA approach. Using Megablast34, we found that the sequence clusters labelled Candida [1], [2] and [4] were predominantly composed of C. albicans, Candida [6] was primarily Candida tropicalis, while Candida [5] had a mixed composition. These results suggest that C. albicans likely displaced the other fungi in the faecal pellets, including other Candida species such as C. tropicalis that were present in the mouse gut before colonization. It was also notable that C. tropicalis was detectable only by ITS sequencing and not by quantitative culture suggesting a possible specific adaptation to the conditions of the GI tract and an inability to grow on the standard fungal medium that was used.
The antibiotic-microbiome-cytokine interface: Antibiotic treatment influenced cytokine mRNA levels primarily through effects on the bacterial and not the fungal microbiota
Our exploratory analysis showed that both antibiotics significantly suppressed cytokine mRNA levels in the terminal ileum. We built BMA linear regression models to identify the underlying changes in microbial genera associated with the altered cytokine expression. In these models, the cytokine mRNA expression level served as the continuous response. The microbiota and categorical constructs representing antibiotic treatment and C. albicans exposure served as the independent covariables. We estimated a separate model for each cytokine.
In the faecal pellets, the only genus with a fairly consistent positive effect on any cytokine was Barnesiella, which was positively associated with IL-22 at day 21 (PIP = 49%) (Figure 5a). In contrast, we observed several consistent effects in the terminal ileum especially on day 7. The genus Clostridium and the related genus, “Candidatus Arthromitus” had the most consistent and significant influence on cytokines. On day 7, Clostridium [2] was positively associated with IL-17A (PIP = 92%) and IL-22 (PIP = 57%) while “Candidatus Arthromitus” was positively associated with IFN-γ (PIP = 78%). Other consistent positive associations in the terminal ileum involved genera of the phylum Proteobacteria, including Phyllobacterium on IL-21 (PIP = 84%), Proteus on IL-17A (PIP = 81%) and Comamonas on IL-21 (PIP = 53%). The only genera with negative cytokine associations were Lactococcus on TNF-α (PIP = 67%) and Veillonella on IL-21 (PIP = 49%). On day 21, the only highly consistent positive effect was that of “Candidatus Arthromitus” on IL-22 (PIP = 96%). In the fungal models (Figure 5b), the only genus with a consistently positive association with IL-21 on day 7 was Phoma in the faecal pellets (PIP = 72%). On day 7, treatment with either PSG or vancomycin had a negative influence on IFN-γ in both GI sites. On day 21, the introduction of C. albicans without concurrent antibiotic treatment was associated with increased IL-17A mRNA levels in both GI sites and with increased IL-22 in the faecal pellets.

Effect of the microbiome and antibiotic treatment on host cytokine mRNA expression.
Results from BMA linear regression ensembles that explain cytokine expression level as a function of the microbiome and antibiotic treatment. A separate model was estimated for each cytokine. Panels (a) and (b) show findings in the bacterial and fungal models, respectively. We show only the effects that were statistically significant i.e. the corresponding Bayesian 95% CIs did not include zero. The effect sizes and the Bayesian 95% CIs of the top consistent and significant variables are presented. A higher value of PIP (expressed in %) indicates that the variable is consistently associated with cytokine expression levels across the space of models explored in BMA.
Overall, both bacterial and fungal genera had relatively consistent positive associations with the panel of cytokines that were studied. Most of these bacteria were depleted due to antibiotic administration, thus explaining the decrease in the overall cytokine levels that we observed in our exploratory analyses (Figure 3b). The negative association of antibiotic treatment with IFN-γ only surfaced in the fungal model on day 7. Similarly, the positive effects of C. albicans on IL-17A and IL-22 only appeared in the fungal model. These differences between the bacterial and fungal models likely represent the stronger effects of the bacteria on the cytokines as compared to the fungi. The strong antibiotic effects on cytokines within the fungal models indicate that the antibiotic variables were proxies for the relevant effects of antibiotics on the bacterial microbiome.
The antibiotic-microbiome-cytokine-colonization interface: Antibiotic-shaped bacterial genera explained levels of C. albicans colonization more consistently than either fungal genera or cytokines
In our final set of multivariable BMA models, we identified the microbiota and cytokines most influential on the level of C. albicans colonization. We built separate BMA linear regression models for bacterial and fungal genera and one for each GI site (Figure 6). The covariables in the models consisted of the microbial genera, cytokine mRNA expression levels, day of sampling, antibiotic treatment and C. albicans exposure. The level of C. albicans colonization (measured as CFUs in the faecal pellets) was the continuous response variable.

Effect of the microbiome, cytokine expression and antibiotic treatment on C. albicans colonization.
Results from BMA linear regression ensembles that explain colonization in log(CFU/g) as a function of the microbiome, cytokine expression levels and antibiotic treatment. Findings from the bacterial models are presented in panels (a) (faecal pellets) and (b) (terminal ileum) while those from the fungal models are presented in panels (c) (faecal pellets) and (d) (terminal ileum). From left to right, each panel consists of (left) a plot of effect size vs. PIP, (middle) 95% Bayesian CI of effect sizes and (right) heatmaps of scaled microbe abundances across control and treatment groups on days 7 and 21. All analyses were performed at the genus level. In the plots showing effect size vs. PIP, each point denotes a variable coloured by its phylum membership or category (e.g. “Experimental”). PIP is expressed in %. A higher value of PIP indicates that the variable is consistently associated with colonization across the space of models explored in BMA. All plots show only the top consistent variables that attained the highest PIPs and whose effects on C. albicans colonization were statistically significant i.e. the corresponding Bayesian 95% CIs did not include zero. The heatmaps show relative abundances (scaled to the range 0–1) of the most influential bacteria and fungi that consistently explained levels of C. albicans colonization across PSG (P), vancomycin (V) and controls (C) groups on days 7 and 21.
In each of the models, only a handful of variables obtained a high PIP (Figure 6). Genera from the phylum Firmicutes had the highest PIP in both the faecal pellets and the terminal ileum (Figure 6a and b). In the faecal pellets, the bacterial genera with the highest PIPs were Streptococcus and Parabacteroides (Figure 6a). Both were positively associated with higher levels of C. albicans colonization and had increased relative abundance in PSG-treated mice. Lactobacillus and Prevotella were protective against colonization, although they had lower PIPs. While the relative abundance of Prevotella was higher in the control mice, the relative abundance of Lactobacillus was higher mainly in the vancomycin-treated mice. In the terminal ileum, Veillonella and to a lesser extent, Enterococcus, were the primary genera positively associated with higher levels of colonization (Figure 6b). Both these genera increased under PSG treatment.
In the fungal models, variables encoding PSG treatment both with and without concomitant C. albicans exposure were the most influential, with a large positive effect on colonization and a PIP of 100% (Figure 6c and d). All the other fungal and cytokine variables had very low PIPs. When this finding is viewed together with the high PIPs assigned to specific bacterial genera in the bacterial model, it is likely that the exclusion of these bacteria in the fungal model resulted in high PIP assignment to the categorical variables for PSG treatment and C. albicans exposure, which served as proxies for the bacterial effects.
Using model diagnostics (see supplementary information for details), we determined that the bacterial model estimated from the faecal pellets was able to explain as much as 90% of the observed variation in C. albicans colonization levels, thus demonstrating good explanatory power. Furthermore, the intercept in the bacterial model had a very low PIP (≈0%) indicating that the model was estimated using a large fraction of the relevant variables influential on C. albicans colonization levels.
The combined findings from the bacterial and fungal models indicate that antibiotic-induced changes in the GI bacterial microbiota and the antibiotics themselves constituted a far more influential effect on colonization than either the resident GI fungi or cytokines.
Discussion
Broad-spectrum antibiotics are known to have a wide-ranging impact on the gut microbiome35 and immunomodulatory effects on both the innate and adaptive components of the immune system19,36. Both the GI flora and the local GI immune response have the potential to prevent or limit GI colonization by C. albicans. In this study, we employed BMA, a Bayesian modelling approach, to examine the complex interface connecting antibiotics, the microbiome, cytokines and C. albicans colonization. Within each facet of this interface, BMA enabled us to identify the most influential variables and rank their relative contributions. Our results indicate that members of the antibiotic-influenced bacterial microbiome had the most consistent and substantial influence on C. albicans colonization than either the fungal microbiome or the local immune response.
Other groups have studied experimental setups that are similar to ours4,5,6,7. However, our experiments differ from these studies in several important aspects. Our primary focus was on obtaining mechanistic insights into C. albicans colonization during a longer span of antibiotic treatment (21 days). In contrast, the earlier studies examined either reassembly of the microbiome after discontinuation of a short duration of antibiotics in the presence and absence of C. albicans exposure4,5,6, or the impact of very long-term antibiotics without a specific focus on C. albicans colonization7. While these studies primarily examined the differential abundance of microbiota across treatments, our modelling framework extends these analyses to simultaneously examine the impact of long-term antibiotics on the gut microbiome and the host immune response and their combined influence on C. albicans colonization. Furthermore, we studied colonization as a quantitative response by contrasting the effects of two antibiotic treatments (vancomycin and PSG) that induce differential levels of C. albicans colonization.
Mason et al.5 showed that introducing C. albicans prevents Lactobacillus species from repopulating the GI tract post-antibiotics and promotes the growth of Enterococcus faecalis. We complement their findings by showing that Lactobacillus is associated with protection against C. albicans colonization in the faecal pellets, while Enterococcus in the terminal ileum is positively associated with C. albicans colonization. We also found that treatment with vancomycin was associated with enhanced growth of Lactobacillus in the faecal pellets. It is thus tempting to speculate that the increased growth of this inhibitory genus was one of the reasons why mice that received vancomycin had only transient C. albicans colonization.
We further extend these mechanistic insights by showing that in the faecal pellets, Streptococcus and Parabacteroides appeared to promote C. albicans colonization and thus act antagonistically to Lactobacillus and Prevotella. Streptococcus was unique because this genus was consistently associated with colonization but not with either antibiotics or cytokines. This finding suggests that the colonization promoting action of Streptococcus involves pathways that do not involve either the immune response panel that we examined or the differential abundance effects induced by the antibiotics. This ties in well with recent research that suggests that Streptococci and Candida species interact through several molecular mechanisms to promote synergistic infection of the oral mucosa37,38. It is possible that similar mechanisms are prevalent in the GI tract.
In the cytokine models, we identified consistent positive effects of the genus Clostridium on the expression of IL-17A and IL-22 mRNA. The closely related genus “Candidatus Arthromitus” was also found to strongly promote IL-22 and IFN-γ and was the only genus to strongly influence the cytokine response on day 21. These results are in agreement with previous reports of the stimulatory effects of “Candidatus Arthromitus” on the GI immune response8,16,17,18,39,40. Furthermore, we identified several other genera, such as Phyllobacterium, Proteus, Comamonas, Lactococcus and Pandoraea that likely influence GI cytokine mRNA levels in the terminal ileum. While these results need to be verified experimentally, they demonstrate the power of BMA to identify bacteria that may play important roles in shaping the GI immune response.
It was notable that neither Clostridia nor “Candidatus Arthromitus” were associated with C. albicans colonization. Conversely, other bacteria such as Streptococcus in the faecal pellets were strongly associated with C. albicans colonization but not with cytokine mRNA levels. These contrasting results suggest that in the faecal pellets, the bacterial influence on C. albicans colonization was either direct or mediated by other mechanisms that did not involve the host GI immune response. A possible exception is Veillonella, which was associated with both suppression of IL-21 and stimulation of C. albicans colonization in the terminal ileum. Our findings are supported by recent work by Vautier et al.41 who showed that both IL-17 knockout mice and mice treated with an IL-1 blocker had normal levels of C. albicans GI colonization, which suggests that these cytokines may not play a significant role in controlling the level of colonization. Given these interesting and varied associations between the microbiota and host immune response, we expect that analysing an expanded panel of cytokines and immune effector cells could yield further insights into the host factors that govern C. albicans colonization.
Fungi other than C. albicans appeared to be largely displaced by C. albicans colonization and, in addition, were not a substantial influence on the host cytokine expression. Among these displaced fungi were those that mapped to C. tropicalis. However, we could not detect this species using quantitative culture. Our findings are thus in agreement with those reported by Iliev et al.42 and suggest that the strain(s) of C. tropicalis that grow in in the mouse GI tract may not grow under standard culture conditions.
A limitation of the current work is that our models did not incorporate absolute microbiota counts. However, model diagnostics showed that the BMA ensembles were able to explain up to 90% of the observed variation in C. albicans colonization levels. This indicates that changes in the absolute microbiota counts would be unlikely to add substantially to the model explaining C. albicans colonization levels, over and above the explanation afforded by the variables already in the model, including relative abundances of microbiota, cytokine expression levels and other experimental variables. Nevertheless, it is possible that absolute microbiota counts may constitute a more direct influence on colonization by other microorganisms in alternative models.
It is possible to apply frequentist stability-based approaches to explore large model spaces for variable selection in microbiome analysis43. However, the key advantage of BMA is its ability to explore a much larger number of microbiome and cytokine variable configurations. Unlike frequentist approaches, BMA can simultaneously identify the most influential variables, capture heterogeneity in the effects of these variables and provide uncertainty estimates around the effect sizes without any assumptions about their distributions31,44. Employing BMA substantially reduces the likelihood of false positives that typically accumulate in multi-step modelling approaches.
BMA findings suggest that there were many substantial effects of individual members of the microbiome community on C. albicans colonization that were not uniformly strong across the samples. Although a majority of these effects may not be important by themselves, the combination of several such weak effects across the microbiome community likely constituted a substantial influence on C. albicans colonization.
Thus, BMA provided a broader insight into the nature and distribution of effects within the data. Incorporating versatile statistical methods such as BMA in microbiome preclinical studies is a strong step towards identifying consistent biomarkers that have a high likelihood of validation by downstream studies45.
In conclusion, PSG and vancomycin have complex effects on the bacterial microbiome and also significant effects on the host immune response. PSG distinctly favours C. albicans colonization through direct effects on the bacterial microbiome. Although PSG also alters the GI immune response and fungal microbiome, these changes have a lesser influence on the level of C. albicans colonization of the GI tract.
Methods
Mouse experiments
Male C57BL/6 mice (Taconic Farms) were housed in high efficiency particulate air (HEPA) filtered cages with sterilized bedding and fed autoclaved chow and water ad libitum throughout the experiment. Prior to antibiotic treatment, mouse faecal pellets were plated onto Sabouraud agar containing chloramphenicol and streptomycin to verify that the mice were not colonized with Candida spp. The mice were divided into a control and two treatment groups. The following antibiotics were added to the drinking water of mice in the two treatment groups and continued for the duration of the experiment: a) vancomycin (0.5 mg/ml) and b) a combination of penicillin (1.5 mg/ml), streptomycin (2 mg/ml) and gentamicin (0.1 mg/ml) (PSG) (Figure 1). After 7 days of antibiotic treatment, 105 yeast cells of C. albicans SC5314 were administered by gavage to some of the mice. Others were continued on antibiotic treatment alone and were not colonized. All colonized mice were quarantined to prevent cross-infection. The mouse studies were carried out in accordance with the National Institutes of Health guidelines for the ethical treatment of animals. This protocol was approved by the Institutional Animal Care and Use Committee of the Los Angeles Biomedical Research Institute at Harbor-UCLA Medical Center.
Sample collection, DNA and RNA extraction
Experiments were initiated with 10 to 12 mice in each control and treatment group. On days 7, 9, 14 and 21, freshly obtained faecal pellets were weighed, homogenized and then quantitatively cultured on Sabouraud agar containing chloramphenicol and streptomycin to measure the number of C. albicans CFUs. On days 7 and 21, 3 to 5 mice per treatment group were humanely euthanized and segments of their terminal ileum were harvested. A portion of these segments was cut lengthwise and vortexed briefly in PBS to separate the loosely adherent material from the mucosa-associated contents. Microbial DNA was isolated from these contents with the QIAamp DNA Stool Minikit (Qiagen) using the protocol from Wu et al.46 modified to include a bead-beating step for more efficient lysis and improved nucleic acid recovery. Autoclaved mouse chow was also processed to evaluate the contribution of ingested fungal and bacterial DNA. A separate portion of the ileum segments were immersed in RNAlater (Qiagen) immediately after collection and RNA was isolated using the RiboPure RNA Purification Kit (Life Technologies). After preparing cDNA using the RETROscript reverse transcriptase kit (Life Technologies), mRNA expression levels of IL-17A, IL-22, IL-21, TNF-α, IFN-γ and IL-4 were measured by real-time PCR and normalized using GAPDH by the ΔΔCt method. At a minimum, samples and data were available from at least 3–5 mice for each treatment arm and for each experiment. Each experiment was repeated at least twice.
Amplification and sequencing
Primer pairs targeting the V3–V5 region of bacterial 16S rRNA gene and fungal internal transcribed spacer (ITS) region were used to amplify the extracted microbial DNA. All amplified sequences contained 10 bp unique barcodes and 454 FLX Titanium adaptors. The primer sequences and amplification strategy are described on the accompanying website47. The PCR products were cleaned using the Qiagen MinElute 96 UF PCR Purification Kit and quantified using the Invitrogen Quant-iT™ High-Sensitivity DNA Assay Kit. 25 ng of amplified DNA was pooled from each sample, concentrated and cleaned up to remove any residual primers or nucleotides using the Agencourt AMPure XP Kit. The amplicons were sequenced on the 454 FLX Titanium sequencing platform.
Taxonomic classification
Taxonomic classification of the 16S and ITS sequences was performed using YAP, a distributed bioinformatics workflow48, described on the accompanying website47. YAP integrates established software modules for 16S and ITS read processing and performs several quality filtering and trimming steps including removal of ambiguous, very short (<220 bp), chimeric and non-16S sequences using implementations of PyroNoise49 and uchime50 within mothur51. The filtered and trimmed sequences were efficiently clustered at 97% similarity level using CD-HIT52 to produce operational taxonomic units (OTU). These OTUs were taxonomically classified up to the genus level using mothur's implementation of the naive Bayes classifier53. OTUs that remained unclassified at the genus level were assigned taxonomies using the NCBI Megablast algorithm34. For genera of particular interest, for example, Candida, the most predominant species level taxonomic labels were also identified using Megablast34. Predominant species were the ones that were most frequent within a particular genus, belonged to the largest OTU clusters, occurred in the largest number of mice samples and obtained the lowest E-value in Megablast.
16S and ITS Databases
The YAP workflow incorporates high quality databases to verify 16S and ITS origin of sequences, check for chimeras and assign taxonomies. For bacterial 16S, the SILVA database54 and a training dataset from the RDP project55 were used. For fungal ITS, a custom database was built as follows. First, a seed database of the entire fungal ribosomal region consisting of 236 diverse ribosomal sequences was compiled by running NT BLAST on an annotated ribosomal region of Aspergillus fumigatus Af29356. Next, 155,136 unique ITS sequences were retrieved from the NCBI public database, aligned to the seed database and oriented in the 5′ to 3′ direction. Clustering was performed in two iterations. The first, at 100% sequence identity, yielded 149,266 clusters and the second, at 97% sequence identity, yielded 39,548 species level clusters. Annotations were reassigned for 2,153 sequences based on clustering results. Finally, 10,226 sequences taxonomically identified as “uncultured” were queried against the NCBI taxonomy database to obtain the maximum possible taxonomic resolution.
Data processing
After confirming that each of the two independent experimental replicates showed similar trends, sequence count data and the cytokine expression data from these two independent replications were combined. The GAPDH normalized cytokine mRNA expression levels and genera level relative abundances of sequences were log transformed to bring the numerical attributes of the dataset into the same dynamic range.
Exploratory analyses
The 99.6% bootstrap CI of the Welch's test statistic was computed to assess significant differences in microbial diversity and cytokine expression levels between each of the treatment (vancomycin, PSG) and the control groups. Differences were deemed significant when the 99.6% CI did not include zero. The CI was computed using 10,000 bootstrap resamples of the data, was free of distributional assumptions and included a conservative correction for 6 simultaneous hypotheses corresponding to the 6 cytokines. The analyses with the bacterial microbiome was performed using 30 mice; 10 in each treatment group while those with the fungal microbiome was performed on 36 mice; 12 in each treatment group. The computation of the Welch's statistic allowed for unequal variances in the treatment groups.
Multivariable modelling
All multivariable statistical models were estimated using Bayesian Model Averaging (BMA)57 with a spike-and-slab prior distribution58 implemented in the BoomSpikeSlab R package33,59. The spike prior is based on an assumption that a sparse set of variables can explain the response. It consists of a Bernoulli distribution that specifies whether or not a variable is selected as influential. The slab prior is a Gaussian distribution that models the effect sizes of the variables, conditional on their being chosen as influential. BMA combines information from these two priors and uses a Markov Chain Monte Carlo procedure to compute a space of 10,000 likely variable configurations that explain the response58. The median effect size and its associated 95% Bayesian CI for each variable was computed using the distribution of its regression coefficient across the 10,000 models. The Bayesian 95% CI was free of distributional assumptions. The initial 1,000 models were excluded as burn-in59. The posterior inclusion probability (PIP) for each variable is the proportion of the models that selected the variable as influential. In our findings, the median effect size and its 95% Bayesian CI and the PIP are presented as the two formal measures of statistical significance and consistency for each variable.
BMA model specifications
Three separate sets of BMA models were estimated: a) logistic regression with antibiotic treatment (PSG or vancomycin) as response and controls as reference; linear regression with b) log(mRNA cytokine expression) as response and c) colonization levels measured in log(CFU) as response. These models were used to evaluate a) impact of antibiotics on specific microbiota and the immune response, b) influence of specific microbiota on cytokine mRNA expression c) the role of specific microbiota and cytokine mRNA expression on the level of C. albicans colonization. In each model, antibiotic treatment (PSG, vancomycin), exposure to C. albicans (+/− C. albicans gavage) and time-points of sampling (Day 7, 21) were included as indicator variables. Mice in control groups served as the reference baseline. Each bacterial model was estimated from 30 mice, 10 in each treatment group (controls, vancomycin and PSG). Each fungal model was estimated from 36 mice, 12 in each treatment group. All data analyses were performed in the R language for statistical computing60.
Data and code availability
Sequencing data, experimental data and associated documentation are available on the accompanying website for our project47. Source code for our analysis is available at GitHub30.
References
Whittington, A., Gow, N. A. R. & Hube, B. From commensal to pathogen: Candida albicans. Human Fungal Pathogens. Kurzai, O. (ed.) 3–18 (Springer-Verlag, Berlin Heidelberg, 2014).
Koh, A. Y. Gastrointestinal Colonization of Fungi. Curr. Fungal Infect. Rep. 7, 144–151 (2013).
Wisplinghoff, H. et al. Nosocomial bloodstream infections due to Candida spp. in the USA: species distribution, clinical features and antifungal susceptibilities. Int. J. Antimicrob. Agents. 43, 78–81 (2014).
Mason, K. L. et al. Candida albicans and bacterial microbiota interactions in the cecum during recolonization following broad-spectrum antibiotic therapy. Infect. Immun. 80, 3371–3380 (2012).
Mason, K. L. et al. Interplay between the gastric bacterial microbiota and Candidaalbicans during postantibiotic recolonization and gastritis. Infect. Immun. 80, 150–158 (2012).
Erb Downward, J. R., Falkowski, N. R., Mason, K. L., Muraglia, R. & Huffnagle, G. B. Modulation of post-antibiotic bacterial community reassembly and host response by Candidaalbicans. Sci. Rep. 3, 2191 (2013).
Dollive, S. et al. Fungi of the murine gut: episodic variation and proliferation during antibiotic treatment. PLoS One. 8, e71806 (2013).
Kumar, S., Bansal, A., Chakrabarti, A. & Singhi, S. Evaluation of Efficacy of Probiotics in Prevention of Candida Colonization in a PICU-A Randomized Controlled Trial. Crit. Care Med. 41, 565–72 (2013).
Roy, A., Chaudhuri, J., Sarkar, D., Ghosh, P. & Chakraborty, S. Role of Enteric Supplementation of Probiotics on Late-onset Sepsis by Candida species in Preterm Low Birth Weight Neonates: A Randomized, Double Blind, Placebo-controlled Trial. N. Am. J. Med. Sci. 6, 50–57 (2014).
Wagner, R. D. et al. Probiotic effects of feeding heat-killed Lactobacillus acidophilus and Lactobacillus casei to Candidaalbicans-colonized immunodeficient mice. J. Food Prot. 63, 638–644 (2000).
Villena, J., Salva, S., Aguero, G. & Alvarez, S. Immunomodulatory and protective effect of probiotic Lactobacillus casei against Candidaalbicans infection in malnourished mice. Microbiol. Immunol. 55, 434–445 (2011).
Underhill, D. M. & Iliev, I. D. The mycobiota: interactions between commensal fungi and the host immune system. Nat. Rev. Immunol. 14, 405–416 (2014).
Puel, A. et al. Inborn errors of human IL-17 immunity underlie chronic mucocutaneous candidiasis. Curr. Opin. Allergy Clin. Immunol. 12, 616–622 (2012).
Yang, Y. et al. Focused specificity of intestinal TH17 cells towards commensal bacterial antigens. Nature 510, 152–156 (2014).
Atarashi, K. et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331, 337–341 (2011).
Ivanov, I. I. et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 139, 485–498 (2009).
Goto, Y. et al. Segmented filamentous bacteria antigens presented by intestinal dendritic cells drive mucosal Th17 cell differentiation. Immunity 40, 594–607 (2014).
Lécuyer, E. et al. Segmented filamentous bacterium uses secondary and tertiary lymphoid tissues to induce gut IgA and specific T helper 17 cell responses. Immunity 40, 608–620 (2014).
Kwiatkowska, B. et al. Immune system as a new therapeutic target for antibiotics. Adv. Biosci. Biotechnol. 04, 91–101 (2013).
Koh, A. Y., Kohler, J. R., Coggshall, K. T., Van Rooijen, N. & Pier, G. B. Mucosal Damage and Neutropenia Are Required for Candidaalbicans Dissemination. PLoS Pathog. 4, e35 (2008).
Koh, A. Y. Murine models of Candida gastrointestinal colonization and dissemination. Eukaryot. Cell 12, 1416–1422 (2013).
Gu, S. et al. Bacterial community mapping of the mouse gastrointestinal tract. PLoS One 8, e74957 (2013).
Rogers, G. B. et al. Functional divergence in gastrointestinal microbiota in physically-separated genetically identical mice. Sci. Rep. 4, 5437 (2014).
Breiman, L. Statistical Modeling: The Two Cultures (with comments and a rejoinder by the author). Stat. Sci. 16, 199–231 (2001).
Segata, N. et al. Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60 (2011).
Harrell, F. E. Regression Modeling Strategies: With Applications to Linear Models, Logistic Regression and Survival Analysis (Springer Series in Statistics). (Springer Verlag, New York, 2010).
Greenland, S. Invited commentary: variable selection versus shrinkage in the control of multiple confounders. Am. J. Epidemiol. 167, 523–9; Discussion 530–1 (2008).
Viallefont, V., Raftery, A. E. & Richardson, S. Variable selection and Bayesian model averaging in case-control studies. Stat. Med. 20, 3215–3230 (2001).
Wintle, B. A., McCarthy, M. A., Volinsky, C. T. & Kavanagh, R. P. The Use of Bayesian Model Averaging to Better Represent Uncertainty in Ecological Models. Conserv. Biol. 17, 1579–1590 (2003).
Shankar, J. regeval: A systematic evaluation of high-dimensional, ensemble based regression for exploring large model spaces in microbiome analyses, GitHub. http://github.com/openpencil/regeval (2014). Date of access: 18/12/2014.
Hoeting, J. A., Madigan, D., Raftery, A. E. & Volinsky, C. T. Bayesian model averaging: a tutorial (with comments by M. Clyde, David Draper and E. I. George and a rejoinder by the authors). Stat. Sci. 14, 382–417 (1999).
Romani, L. Immune resistance and tolerance to fungi. G. Ital. Dermatol. Venereol. 148, 551–561 (2013).
Scott, S. L. BoomSpikeSlab: MCMC for spike-and-slab regression. The Comprehensive R Archive Network. http://CRAN.R-project.org/package=BoomSpikeSlab (2014). Date of access: 18/12/2014.
Camacho, C. et al. BLAST+: architecture and applications. BMC Bioinformatics 10, 421 (2009).
Rea, M. C. et al. Effect of broad- and narrow-spectrum antimicrobials on Clostridium difficile and microbial diversity in a model of the distal colon. Proc. Natl. Acad. Sci. U. S. A. 108 Suppl 1, 4639–4644 (2011).
Nau, R. & Tauber, S. C. Immunomodulatory Properties of Antibiotics. Curr. Mol. Pharmacol. 1, 68–79 (2008).
Xu, H., Jenkinson, H. F. & Dongari-Bagtzoglou, A. Innocent until proven guilty: mechanisms and roles of Streptococcus-Candida interactions in oral health and disease. Mol. Oral Microbiol. 29, 99–116 (2014).
Diaz, P. I., Strausbaugh, L. D. & Dongari-Bagtzoglou, A. Fungal-bacterial interactions and their relevance to oral health: linking the clinic and the bench. Front. Cell. Infect. Microbiol. 4 (2014).
Ivanov, I. I. & Honda, K. Intestinal commensal microbes as immune modulators. Cell Host Microbe 12, 496–508 (2012).
Lee, Y. K. & Mazmanian, S. K. Microbial learning lessons: SFB educate the immune system. Immunity 40, 457–459 (2014).
Vautier, S. et al. Candida albicans colonization and dissemination from the murine gastrointestinal tract: the influence of morphology and Th17 immunity. Cell Microbiol. 10.1111/cmi.12388 (2014).
Iliev, I. D. et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 336, 1314–1317 (2012).
Lin, W., Shi, P., Feng, R. & Li, H. Variable selection in regression with compositional covariates. Biometrika. 10.1093/biomet/asu031 (2014).
Scott, S. L. Data augmentation, frequentist estimation and the Bayesian analysis of multinomial logit models. Statist. Papers 52, 87–109 (2011).
Perrin, S. Preclinical research: Make mouse studies work. Nature 507, 423–425 (2014).
Wu, G. D. et al. Sampling and pyrosequencing methods for characterizing bacterial communities in the human gut using 16S sequence tags. BMC Microbiol. 10, 206 (2010).
Shankar, J. bayesianmice: A Bayesian model-based investigation of Candida albicans colonization in a preclinical mouse model, GitHub. http://openpencil.github.io/bayesianmice (2014). Date of access: 18/12/2014.
Szpakowski, S. YAP: A Computationally Efficient Workflow for Taxonomic Analyses of Bacterial 16S and Fungal ITS Sequences, GitHub. http://github.com/shpakoo/YAP (2013). Date of access: 18/12/2014.
Schloss, P. D., Gevers, D. & Westcott, S. L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS One 6, e27310 (2011).
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. & Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200 (2011).
Schloss, P. D. et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541 (2009).
Huang, Y., Niu, B., Gao, Y., Fu, L. & Li, W. CD-HIT Suite: a web server for clustering and comparing biological sequences. Bioinformatics 26, 680–682 (2010).
Wang, Q., Garrity, G. M., Tiedje, J. M. & Cole, J. R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 73, 5261–5267 (2007).
Quast, C. et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596 (2013).
Cole, J. R. et al. The Ribosomal Database Project: improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 37, D141–D145 (2009).
Arnaud, M. B. et al. The Aspergillus Genome Database (AspGD): recent developments in comprehensive multispecies curation, comparative genomics and community resources. Nucleic Acids Res. 40, D653–D659 (2012).
Hoeting, J. A., Madigan, D., Raftery, A. E. & Volinsky, C. T. Bayesian Model Averaging: A Tutorial. Stat. Sci. 14, 382–417 (1999).
George, E. I. & Mcculloch, R. E. Approaches for Bayesian variable selection. Statistica Sinica 7, 339–373 (1997).
Scott, S. L. & Varian, H. R. Predicting the Present with Bayesian Structural Time Series. IJMNO 5, 4–23 (2014).
R Development Core Team. R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing. http://www.R-project.org (2014). Date of access: 18/12/2014.
Acknowledgements
We thank the authors of BoomSpikeSlab, scales, ggplot2, plyr, reshape2, data.table, Hmisc, RcolorBrewer, R Studio and the R Core Development Team for their statistical packages. We thank Benjamin Rosen for his constructive comments and Reed Shabman for contributing “Reed's mouse” to our manuscript. We gratefully acknowledge the collaborative platforms from Google Docs, Paperpile and GitHub. This work was supported by NIH grants R01AI054928 and R01DE017088 and OPP1017579 from the Bill and Melinda Gates Foundation.
Author information
Authors and Affiliations
Contributions
J.S. and S.G.F. interpreted the analyses and wrote the manuscript. J.S. designed and implemented the statistical analysis. N.V.S. and H.L. conducted the mouse experiments and DNA/RNA extractions. S.M. conducted the sequencing experiments. S.S. designed and implemented the bioinformatics workflow. S.G.F., W.C.N., N.V.S., S.M. and L.L. designed the study. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Shankar, J., Solis, N., Mounaud, S. et al. Using Bayesian modelling to investigate factors governing antibiotic-induced Candida albicans colonization of the GI tract. Sci Rep 5, 8131 (2015). https://doi.org/10.1038/srep08131
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep08131
This article is cited by
-
Epidemiology, Clinical Characteristics and Risk Factors for Severity of Chronic Disseminated Candidiasis in Jerusalem, Israel
Mycopathologia (2023)
-
Temporal changes in gastrointestinal fungi and the risk of autoimmunity during early childhood: the TEDDY study
Nature Communications (2022)
-
Dairy associations for the targeted control of opportunistic Candida
World Journal of Microbiology and Biotechnology (2021)
-
Adaptation of Candida albicans During Gastrointestinal Tract Colonization
Current Clinical Microbiology Reports (2018)
-
Infant fungal communities: current knowledge and research opportunities
BMC Medicine (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
