
Abstract
The Eocene–Oligocene Boundary (~34 million years ago) marks one of the largest extinctions of marine invertebrates in the world oceans and of mammalian fauna in Europe and Asia in the Cenozoic era. A shift to a cooler climate across this boundary has been suggested as the cause of this extinction in the marine environment, but there is no manifold evidence for a synchronous turnover of flora, fauna and climate at the Eocene–Oligocene Boundary in a single terrestrial site in Asia to support this hypothesis. Here we report new data of magnetostratigraphy, pollen and climatic proxies in the Asian interior across the Eocene–Oligocene Boundary; our results show that climate change forced a turnover of flora and fauna, suggesting there was a change from large-size perissodactyl-dominant fauna in forests under a warm-temperate climate to small rodent/lagomorph-dominant fauna in forest-steppe in a dry-temperate climate across the Eocene–Oligocene Boundary. These data provide a new terrestrial record for this significant Cenozoic environmental event.
Similar content being viewed by others
Introduction
The Eocene–Oligocene Boundary was one of the most pronounced climatic events of the Cenozoic era, marking a dramatic shift from a “greenhouse” to an “icehouse” world1,2,3,4,5,6,7,8. This event also characterizes one of the largest mammalian fauna turnovers and extinctions, named the Grande Coupure in Europe4,9. In Asia, Meng and McKenna5 reviewed fossil records from tens of sites in Mongolia and northern China, which yielded a composite record showing a turnover in the fauna named “Mongolian Remodelling”. In addition to the abrupt changes of mammalian fauna, climatic changes have also been identified across the Eocene–Oligocene Boundary10,11,12,13,14. However, there still lacks in situ multidisciplinary studies of flora, fauna and climate focused on a same site in Asia. Here, we present mammalian, vegetation and climatic records from Paleogene deposits that show a synchronous turnover of flora, fauna and climate at the Eocene–Oligocene Boundary.
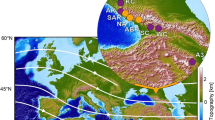
Our study site lies in the Junggar Basin in northwestern China. This basin is situated between the Altay Mountains to the north and the Tianshan Mountains to the south (Fig. 1a). Our section is located at Keziletuogayi (47°50′26″N, 86°40′45″E, 520 m above sea level), adjacent to the Ertix River (Fig. 1b). In the Paleogene deposits there are Late Eocene to Early Oligocene mammalian fossils15,16,17,18,19. However, to date, there has been no study of the vegetation and climatic changes across the Eocene–Oligocene Boundary at this site.

Map showing the Junggar Basin and the location of the studied section in China.
(a), Location of the Junggar Basin. (b), Geological map and the location of the Keziletuogayi section. (Maps was drawn by Jimin Sun using the Software of Canvas 8.0.2).
In order to understand in situ floral, faunal and climatic changes, we collected multiple samples, focusing on the middle to upper parts of the Keziletuogayi section (Fig. 2a). In general, the sedimentary facies are fluvial to lacustrine; the lower part is dominated by bright brown sandstone/siltstone with intercalated white cross-bedded sandstones, while the upper part is characterized by light-yellow sandstone/siltstone (Fig. 2b). Oriented specimens were collected for paleomagnetic polarity studies and bulk samples for analysis of paleovegetational and climatic changes.

Lithology and the magnetostratigraphy of the Keziletuogayi section.
(a), The lithology of the section. (b), Photo showing the boundary (indicated by the red arrow) of the Late Eocene and Early Oligocene strata. (c), Magnetostratigraphy and mammalian fossils zones (A1 to A3) of the Keziletuogayi section. (Photo was taken by Jimin Sun).
The chronology of the studied section is based both on the new magnetostratigraphic data (Supplementary Table 1) and biostratigraphic age control (Fig. 2c). Firstly, based on the previous paleontological studies together with our newly discovered mammalian fossils (Supplementary Table 2), three mammal assemblages (A1 to A3) were identified15,17,18. These mammal assemblages correlate well with Asian and European land mammal ages16,17,18. Among them, A3 corresponds to the Latest Eocene Ulangochuan age (Chinese Land Mammal Ages, CLMA), whereas A2 and A1 correspond to the Early Oligocene Ulantatalian age (Fig. 3a). With the above biostratigraphic age controls, the measured paleomagnetic polarity of the section could be correlated well with the geological time scale 2004 (GTS 2004, Gradstein et al.20) that covers polarity chrons from the upper C16n.2n to the lower C12r and spanning a time range of 36–33 Myr ago (Fig. 2c).

Fauna and Flora records from the Latest Eocene to the Early Oligocene in the Keziletuogayi section.
(a), Mammalian faunas of the section, noting the distinct mammalian turnover across the Eocene–Oligocene Boundary, CLMA: Chinese Land Mammal Ages; ALMA: Asian Land Mammal Ages. (b), Vegetation changed from forests to forest-steppe environment after the beginning of the Oligocene.
Mammalian fossils from A3 consist both of large size perissodactyl mammals and the newly discovered small mammalian fossils, but they are dominated by giant perissodactyl mammals (Fig. 3a). This assemblage is similar to that of the Ergilin fauna of Mongolia5,17, which has a Latest Eocene age. In contrast to the A3 mammal fossils, fossil assemblages of A1 and A2 are dominated by small rodents and lagomorphs (Fig. 3a), comparable to those of the Early Hsandagolian fauna of Mongolia18,21,22,23, which suggests an Early Oligocene age. Therefore, the mammalian fossils of this section reveal a distinct faunal turnover pattern characterized by a shift from Late Eocene, giant perissodactyl-dominant fauna to Early Oligocene, small rodent/lagomorph-dominant fauna (Fig. 3a).
Vegetational changes in the section were reconstructed with pollen analysis (Supplementary Table 3). Our results indicate two distinct vegetation types across the Eocene–Oligocene Boundary (Fig. 3b). The Late Eocene terrestrial flora were dominated by broad-leaved species of Betulaepollenites, Quercoidites, Ulmipollenites, Juglanspollenites and coniferous species of Piceaepollenites, Pinuspollenites, Abiespollenites and Podocarpidites, whereas the herb taxa only occupied less than 7 percent (Fig. 3b). The vegetation type during the Late Eocene was a mixed coniferous–broad-leaved forest. However, there was a remarkable change of pollen transitioning into the Early Oligocene (Fig. 3b). The Early Oligocene pollen flora are characterized by an abrupt increase in herb taxa including Artemisiaepollenites, Chenopodipollis, Graminidites, Ephedripites, Rosapollenites, Compositoipollenites and other herb species, whereas the tree species of Juglanspollenites, Quercoidites and Podocarpidites largely decrease, implying a forest-steppe environment with open grasslands.
Further analysis of the pollen species can be used to infer both temperature and aridity changes. The relatively high percentage of mesothermic species of Juglanspollenites and Quercoidites, together with the evergreen hydrothermal species of Podocarpidites suggest a warm-humid climate in the Late Eocene, but such species decreased to lower percentages after the beginning of the Oligocene, suggesting a change to a cooler climate (Fig. 3b). Among the herb taxa, Artemisiaepollenites and Chenopodipollis are very commonly in dry steppes, while Ephedripites is a dry shrub. The abrupt increase in such xerophytic elements suggests that the climate tended to become drier after the start of the Oligocene (Fig. 3b). Therefore, our pollen results reveal an abrupt shift in the ecosystem from warm-humid forest in the Late Eocene to dry-temperate forest-steppe in the Early Oligocene.
Recent reviews of Paleogene palynoflora in eastern China also revealed evidence of cooling from the Late Eocene into the Oligocene24, but lack evidence for a drier climate. A possible cause for a wet climate in the Oligocene in eastern China is the short distance to the Pacific Ocean compared with the remote inland basins in western China.
Therefore, the enhanced cooling and aridity indicated by the pollen data in the Junggar Basin was essentially synchronous with the beginning of the Oligocene and it is also consistent with the turnover time of mammals. Similar results from the Hampshire Basin of England show that faunal turnover in the rodents in the latest pre-Grande Coupure fauna coincided with a change in vegetation types9.
Our new paleoclimatic reconstructions are based on multiple climatic proxy parameters (Supplementary Table 4) (Fig. 4). Clay minerals are very common in sedimentary rocks and soils; their categories and the relative concentrations of different minerals are mostly dependent on the degree of chemical weathering. More specifically, kaolinite forms in warm and humid climates, particularly under tropical and subtropical conditions25. Fig. 4a clearly indicates that the content of kaolinite is much higher in the Late Eocene sediments, but it decreases abruptly above the Eocene–Oligocene Boundary, suggesting weaker chemical weathering and a relatively cool-dry climate after the beginning of the Oligocene. Recent mineralogical studies of Cenozoic sediments in the Tarim Basin, which is a neighboring basin, located to the south of the studied Junggar Basin, also indicate a dramatic decrease of kaolinite from the Late Eocene to Early Oligocene, implying climatic cooling at and after the end of the Eocene26.

Paleoclimatic changes indicated by various climatic proxies.
(a), Variations of kaolinite versus depth. (b), Vertical fluctuations of color index of redness (a*). (c), Variations of Fe2O3 content of the Keziletuogayi section. (d) and (e), High-resolution curves of the redness (a*) and the Fe2O3 content across the Eocene–Oligocene Boundary. Red arrow indicates the variation trend at the Eocene–Oligocene Boundary.
Sediment color has long been used as a paleoclimatic parameter27. The redness (a*) is related to the presence of reddish crystalline iron oxides such as goethite and hematite that form during chemical weathering and accordingly the redness is climate dependent27. Fig. 4b shows that there is a general trend towards lower values, suggesting there was weakening chemical weathering after the start of the Oligocene.
Due to the extremely low solubility of Fe oxides over the normal pH range of soils, Fe2O3 produced during mineral weathering is precipitated mainly as hematite and goethite. The content of Fe2O3 has been used as a parameter in reconstructing past climatic change28. Because the iron oxides have a pronounced effect on sediment color, this can account for the very similar variation trends between the content of Fe2O3 and the redness (Fig. 4b, c). A similar relationship was reported in the long-term paleoclimatic changes in the Qaidam Basin28, which is also a large inland basin in the northwest China. The higher values of both redness and Fe2O3 content imply stronger chemical weathering under relatively warmer and more humid climatic conditions during the Eocene. The climatic shift at the Eocene–Oligocene Boundary is more remarkable in the high-resolution curve across the boundary (Fig. 4d, e).
Therefore, our new data from the interior of Asia demonstrate simultaneous changes in fauna, ecosystem and climate, suggesting a change from a perissodactyl-dominant fauna in forests in a warm-temperate climate in the Late Eocene to a small rodent/lagomorph-dominant fauna in a forest-steppe, dry-temperate climate in the Early Oligocene (Supplementary Figure 1); this provides an important new terrestrial record of dramatic environmental changes at the Eocene–Oligocene Boundary.
Moreover, it has become increasingly apparent that Earth's climate underwent a dramatic change when a permanent ice sheet began to form in Antarctica about 34 million years ago, at the Eocene–Oligocene Boundary8,29. The large ice sheet actively interacted with the rest of the climate system by amplifying, pacing and potentially driving global climatic cooling and this caused a dramatic response in the biosphere. The new record presented here documents important synchronous continental climatic and biotic changes during the transition from the warm Eocene world to the cool Oligocene and therefore has implications for the effect of onset of Antarctic glaciation on regional aridity and the evolution of flora and fauna in Asia. Apart from the Junggar Basin, this aridification since the beginning of the Oligocene is also well documented by several paleoenvironmental studies in the Xining and Tarim Basins as well as in the southern Mongolia in the central Asian continent11,14,26, although Bosboom et al.30 argued that there may be a major hiatus across the Eocene–Oligocene Boundary in the Tarim Basin.
It is interesting to discuss the effects of Tibetan Plateau uplift and global climatic cooling on the paleoenvironmental shift at the Eocene–Oligocene Boundary. Although the Tibetan Plateau uplift was responsible for aridification in central Asia by creating a rain shadow, the timing of the uplift may have been diachronous31. The oblique subduction of the Indian lithosphere beneath Tibet accounts for the stepwise rise of different parts of the Tibetan Plateau ranging from 55 Myr ago in the southwest to the latest uplift of its northeastern margin in the last few million years32,33. To date, there is still a lack of evidence for dramatic uplift of Tibet at 34 Myr ago and therefore there is some uncertainty that the growth of the plateau could have reached a threshold elevation that could trigger a change in atmospheric circulation and climatic patterns. Therefore, the contemporaneity of the coeval changes in climate, plant and animal communities and the initiation of the Antarctic glaciation further demonstrate that the cause of such a synchronous turnover was global cooling rather than regional tectonic uplift.
Methods
Magnetostratigraphy
Specimens for magnetostratigraphic analysis were oriented with a magnetic compass in the field. All samples were subjected to stepwise thermal demagnetization, commonly using the following stepwise heating routine: (i) 100°C or 50°C steps up to 500°C; and (ii) 25°C or 10°C steps up to 640°C or 690°C. The magnetic remanence was measured with a 2G, three-axis, cryogenic magnetometer housed in field-free space (300 nT) at the Institute of Geology and Geophysics, Chinese Academy of Sciences. Thermal demagnetization of the samples revealed two remanence components: a secondary magnetization with low unblocking temperature; and a characteristic remanent magnetizations (ChRM) with high unblocking temperatures mostly between 450°C and 670°C. The stable ChRM direction of each specimen was calculated using linear regression techniques and the mean site value was obtained following Fisher's statistics.
Pollen analysis
At least 100 g of sediment was used for each sample in order to acquire enough pollen grains. Samples were treated with HCl (35%) and HF (70%) to remove carbonates and silica. Separation of the palynomorphs from the residue was performed by using ZnCl2 (density = 2). Slides were prepared by mounting the pollen grains in glycerin jelly and then counted under a microscope using ×400 magnifications. For each sample, pollen grains were counted until a minimum pollen sum of 200 grains was reached.
Color measurement
For color measurement, samples (1.5 g) were first dried at 40°C for 24 h, then crushed to powder and finally measured using a handheld Minolta-CM2002 spectrophotometer. The color reflectance of all samples are given by the spherical L*a*b* color space, among them, a* represents redness.
Clay mineral analysis
Samples for clay mineral analysis were firstly treated with acetic acid (10%) and H2O2 (30%) to remove carbonates and organic matter. Clay fraction of <2 μm was extracted, pipetting according to Stokes' Law. Mineralogical analyses were completed with a X-ray Diffractometer (DMAX 2400). We estimated that the analytical uncertainties were about ±10%.
Fe2O3 content measurement
For each sample, 0.05 g powder was first acid-treated (5 ml HF, 1 ml HNO3 and 0.5 ml HClO4) and heated, then the treated samples were dissolved in 10 ml HCl and measured using atomic absorption spectroscopy (PinAAcle 900F). The analytical uncertainties were less than about ±5%.
References
Miller, K. G., Fairbanks, R. G. & Mountain, G. S. Tertiary oxygen isotope synthesis, sea level history and continental margin erosion. Paleoceanogr. 2, 1–19 (1987).
Prothero, D. R. & Heaton, T. H. Faunal stability during the early Oligocene climatic crash. Palaeogeogr. Palaeoclimatol. Palaeoecol. 127, 257–283 (1996).
Houben, A. J. P. et al. Reorganization of Southern Ocean Plankton Ecosystem at the Onset of Antarctic Glaciation. Science 340, 341–344 (2013).
Stehlen, H. G. Remarques sur les faunules de Mammifères des couches eocenes et oligocenes du Bassin de Paris. Bull. Soc. Geol. Fr. 9, 488–520 (1909).
Meng, J. & McKenna, M. C. Faunal turnovers of Palaeogene mammals from the Mongolian Plateau. Nature 394, 364–367 (1998).
Zanazzi, A., Kohn, M. J., MacFadden, B. J. & Terry, D. O. Large temperature drop across the Eocene-Oligocene transition in central North America. Nature 445, 639–642 (2007).
Lear, C. H., Elderfield, H. & Wilson, P. A. Cenozoic deep-sea temperatures and global ice volumes from Mg/Ca in benthic foraminiferal calcite. Science 287, 269–272 (2000).
Liu, Z. H. et al. Global cooling during the Eocene- Oligocene climate transition. Science 323, 1187–1190 (2009).
Hooker, J. J., Collinson, M. E. & Sille, N. P. Eocene-Oligocene mammalian faunal turnover in the Hampshire Basin, UK: calibration to the global time scale and the major cooling event. J. Geol. Soc. Lond. 161, 161–172 (2004).
Ivany, L. C., Patterson, W. P. & Lohmann, K. C. Cooler winters as a possible cause of mass extinctions at the Eocene/Oligocene boundary. Nature 407, 887–890 (2000).
Dupont-Nivet, G. et al. Tibetan Plateau Aridification linked to global cooling at the Eocene-Oligocene transition. Nature 445, 635–638 (2007).
Norris, G. Spore-pollen evidence for early Oligocene high-latitude cool climatic episode in northern Canada. Nature 298, 387–389 (1982).
Jaramillo, C., Rueda, M. & Mora, G. Cenozoic Plant Diversity in the Neotropics. Science 311, 1893–1896 (2006).
Kraatz, B. P. & Geisler, J. H. Eocene–Oligocene transition in Central Asia and its effects on mammalian evolution. Geology 38, 111–114 (2010).
Ye, J., Meng, J., Wu, W. Y. & Wu, S. Y. The discovery of late Eocene mammal fossils from Burqin of Xinjiang. Vert. PalAs. 40, 203–210 (2002).
Wu, W. Y., Meng, J., Ye, J. & Ni, X. J. Propalaeocastor (Rodentia, Mammalia) from the Early Oligocene of Burqin Basin, Xinjiang. Am. Mus. Novit. 3461, 1–16 (2004).
Ye, J., Meng, J., Wu, W. Y. & Ni, X. J. Late Eocene–early Oligocene lithological and biological stratigraphy in the Burqin region of Xinjiang. Vert. PalAs. 43, 49–60 (2005).
Ni, X., Meng, J., Wu, W. & Ye, J. A new Early Oligocene peradectine marsupial (Mammalia) from the Burqin region of Xinjiang, China. Naturwiss. 94, 237–241 (2007).
Zhang, R., Kravchinsky, V. A. & Yue, L. Link between global cooling and mammalian transformation across the Eocene–Oligocene boundary in the continental interior of Asia. Int. J. Earth Sci. 101, 2193–2200 (2012).
Gradstein, F., Ogg, J. & Smith. A. A Geologic Time Scale 2004 (Cambridge University Press, Cambridge, 2004).
Höck, V. et al. Oligocene–Miocene sediments, fossils and basalts from the Valley of Lakes (Central Mongolia)— an integrated study. Mitt. Österr. Geol. Ges. 90, 83–125 (1999).
Daxner-Höck, G. Ulaancricetodon badamae n. gen. n. sp. (Mammalia, Rodentia, Cricetidae) from the Valley of Lakes in Central Mongolia. Paläont. Zeitschr. 74, 215–225 (2000).
Daxner-Höck, G. New zapodids (Rodentia) from Oligocene–Miocene deposits in Mongolia. Part 1. Senck. Leth. 81, 359–389 (2001).
Ma, X. L., Jiang, H. C., Cheng, J. & Xu, H. Y. Spatiotemporal evolution of Paleogene palynoflora in China and its implication for development of the extensional basins in East China. Rev. Palaeobot. Palynol. 184, 24–25 (2012).
Islam, M. R., Stuart, R., Risto, A. & Vesa, P. Mineralogical changes during intense chemical weathering of sedimentary rocks in Bangladesh. J. Asian Earth Sci. 20, 889–901 (2002).
Wang, C. W. et al. The Eocene–Oligocene climate transition in the Tarim Basin, Northwest China: Evidence from clay mineralogy. Appl. Clay Sci. 74, 10–19 (2013).
Giosan, L., Flood, R. D. & Aller, R. C. Paleoceanographic significance of sediment color on western North Atlantic drifts: I. Origin of color. Mar. Geol. 189, 25–41 (2002).
Song, B. W., Zhang, K. X., Lu, J. F., Wang, C. W. & Xu, Y. D. The middle Eocene to early Miocene integrated sedimentary record in the Qaidam Basin and its implications for paleoclimate and early Tibetan Plateau uplift. Can. J. Earth Sci. 50, 183–196 (2013).
Zachos, J., Pagani, M., Sloan, L., Thomas, E. & Billups, K. Trends, rhythms and aberrations in global climate 65 Ma to present. Science 292, 686–693 (2001).
Bosboom, R. et al. LinkingTarim Basin sea retreat (west China) and Asian aridification in the late Eocene. Basin Res. 26, 1–20 (2013).
Chung, S. L. et al. Diachronous uplift of the Tibetan plateau starting 40 Myr ago. Nature 394, 769–773 (1998).
Tapponnier, P. et al. Oblique stepwise rise and growth of the Tibet Plateau. Science 294, 1671–1677 (2001).
Rowley, D. B. & Garzione, C. N. Stable isotope-based paleoaltimetry. Ann. Rev. Earth Planet. Sci. 35, 463–508 (2007).
Acknowledgements
This work was supported by the “Strategic Priority Research Program” of the Chinese Academy of Science (XDB03020500), the National Basic Research Program of China (2013CB956400) and the National Nature Science Foundation of China (41290251 and 41272203). We thank Pan, Y.S. and Zhang, Z.Y. for their help in field sampling.
Author information
Authors and Affiliations
Contributions
J.S. wrote the manuscript, X.N., J.Y., W.W., S.B. and J.M. collected and identified mammal fossils. J.S. completed magnetostratigraphic and paleoenvironmental work. B.W. improved the language and suggested for figure revisions. All authors discussed and contributed to the publication.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplelentary Information
Supplementary File
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Sun, J., Ni, X., Bi, S. et al. Synchronous turnover of flora, fauna and climate at the Eocene–Oligocene Boundary in Asia. Sci Rep 4, 7463 (2014). https://doi.org/10.1038/srep07463
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep07463
This article is cited by
-
A punctuated equilibrium analysis of the climate evolution of cenozoic exhibits a hierarchy of abrupt transitions
Scientific Reports (2023)
-
Phylogenomic analyses highlight innovation and introgression in the continental radiations of Fagaceae across the Northern Hemisphere
Nature Communications (2022)
-
Relationship between tectonism and desertification inferred from provenance and lithofacies changes in the Cenozoic terrestrial sequence of the southwestern Tarim Basin
Progress in Earth and Planetary Science (2021)
-
Widespread loss of mammalian lineage and dietary diversity in the early Oligocene of Afro-Arabia
Communications Biology (2021)
-
East Asian paleoclimate change in the Weihe Basin (central China) since the middle Eocene revealed by clay mineral analysis
Science China Earth Sciences (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
