
Abstract
Over the last century humans have altered the export of fluvial materials leading to significant changes in morphology, chemistry and biology of the coastal ocean. Here we present sedimentary, paleoenvironmental and paleogenetic evidence to show that the Black Sea, a nearly enclosed marine basin, was affected by land use long before the changes of the Industrial Era. Although watershed hydroclimate was spatially and temporally variable over the last ~3000 years, surface salinity dropped systematically in the Black Sea. Sediment loads delivered by Danube River, the main tributary of the Black Sea, significantly increased as land use intensified in the last two millennia, which led to a rapid expansion of its delta. Lastly, proliferation of diatoms and dinoflagellates over the last five to six centuries, when intensive deforestation occurred in Eastern Europe, points to an anthropogenic pulse of river-borne nutrients that radically transformed the food web structure in the Black Sea.
Similar content being viewed by others

Introduction
The development of human civilization is the most significant environmental event of the last 10,000 years4. The global impact of human activities has come into focus as fossil fuel-based industrialization has accelerated over the last century, but the expansion of agriculture may have affected the entire Earth System as early as 8000 years ago5. In the ocean, anthropogenic impacts include increasing temperatures, sea level rise, eutrophication and acidification6. Evidence for long term, pre-industrial anthropogenic modifications of the oceans remains sparse1,7,8, although humans became active agents of continental denudation thousands of years ago9. However, the ability to define baselines and separate natural variability from human-induced changes is essential for understanding biogeochemical cycles and managing marine ecosystems, which ultimately depends on the detection and attribution of long term environmental trends6.
Regional seas10 provide the best opportunity to examine early anthropogenic changes in the marine environment. Their amplified reactivity to continental processes is the result of the limited water exchange with the open ocean and the relatively large fluvial contribution to their overall water budget. We selected the Black Sea to examine if pre-industrial land use was recorded by sedimentation patterns and sedimentary biogeochemistry (Fig. 1A). The landlocked Black Sea is a sensitive recorder of continental signals as demonstrated by drastic and repeated alteration of its ecosystem over the past half century2,11. Although evaporation exceeds precipitation, the freshwater budget of the Black Sea is positive due to river runoff12. Low salinity water is exported at the surface toward the Mediterranean through the narrow and shallow Straits of Bosporus; denser water from the Sea of Marmara flows into the Black Sea as an undercurrent. River runoff drives much of the seasonal and interannual variability in salinity and sea level, which in turn affects the water exchange through the Bosporus13. The Danube River, draining ~30% of central and eastern Europe (Fig. 1A), provides over 60% of the entire runoff reaching the Black Sea12. Because fluvial nutrients feed the surface mixed layer that accounts for only 13% of the entire basin volume, there is a strong coupling between Danube fluxes and the Black Sea's biogeochemistry and ecology2,11. A vigorous cyclonic mean surface circulation with superimposed mesoscale eddies13,14 redistributes nutrients in the surface mixed layer at multiannual time scales13, whereas nutrient exchanges with deeper suboxic and anoxic layers are secondary in importance13.

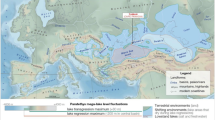
Location of Danube-Black Sea system and the evolution phases of Danube delta.
(A) Black Sea and its watershed (contoured with dashed white line), Danube's watershed (contoured with white line); Danube and Danube delta (blue line and green fill respectively). Location of core GGC18/MC19 is indicated. (B) Geologic map of the Danube delta showing the extent of the present delta plain, the thickness of its deposits (red isopachs in meters) as well as the mapped extent of individual lobes15 (see also SI): Tulcea (pre-5500 yrs BP to Present); St. George I (~5500–3600 yrs BP); Sulina (~3600–2000 yrs BP); St. George II and Dunavatz, (both ~2000 yrs BP to present); Chilia I (older than 1000 yrs BP ); Chilia II (~1000–300 yrs BP); and Chilia III (~300 yrs BP to present). (C) Satellite image showing the expansion of Danube delta over the last 2000 years (land built during this period highlighted with a yellow mask).
After the 1960s, the Black Sea was transformed within a decade into the largest eutrophic water body in the world, by discharge of industrial, domestic and agricultural waste coming primarily from the Danube watershed11. In addition, construction of dams within the Danube watershed substantially reduced dissolved silicate loads to the Black Sea2 hindering siliceous phytoplankton such as diatoms in favor of algal bloom-forming flagellates and coccolithophores. Breakdown of communist economies in Eastern Europe after 1990 led to a partial recovery of Black Sea ecosystems. The eutrophication period, characterized by drastic changes of the food web structure and collapse of fisheries, is illustrative of the sensitivity of the basin. Similarly, it has been postulated that the Danube-Black Sea system was influenced by changes in pre-industrial land use that modulated material fluxes to the Black Sea14.
Results
Danube River was one of the main advance routes for Neolithic agriculture from the Fertile Crescent into Europe and land use changes occurred in its basin as early as 7,500 yrs BP. To reconstruct the history of continental denudation in the Danube basin that accompanied these changes, we generated sequential sediment budgets for the Danube delta. The modern delta covers ca. 4080 km2 and has been built in several phases as bayhead, lacustrine and open-coast delta lobes (Fig. 1B)15. The base of the modern delta is marked by the maximum flooding surface above the previous early Holocene delta that was built during the lake phase of the Black Sea and drowned after the lake reconnected to the ocean at ca. 9000 yrs BP (see SI). Using modern relief as the upper surface, we estimated that ~215 GT of sediments have been stored in the delta above the maximum flooding surface (Fig. 1B). This value corresponds to ~26 MT yr−1 and is considerably lower than Danube's sediment load of ~70 MT yr−1 estimated prior to 1960's, before the most intensive dam construction phase along its course and tributaries16. However, consistent with the higher sediment load estimate, the Danube delta, expanded faster in its later history, which suggests a link to the intensification of land use in the watershed as the population increased and land use technology advanced17.
Radiocarbon and optically stimulated luminescence15 (SI) show that ~40% of the delta plain area was constructed over the last two millennia (Fig. 1C). To quantify this late Holocene acceleration, we assessed the sediment storage corresponding to discrete developmental phases showing that average storage rate increased over five times from the early to late Holocene (Fig. 2A). Higher fluvial sediment discharge is the most likely cause for these increasing deltaic storage rates as the delta front advanced on the shelf. We can exclude enhanced trapping by the delta in its later development phases because the degree of exposure to wave reworking intensified toward the late Holocene as delta lobes advanced outside the sheltered Danube Bay into progressively deeper waters15. Equally so, diminished trapping along the alluvial valley and tributaries upstream of the delta can be discounted because these settings are still underfilled, providing ample accommodation space for sediments (SI). Our storage rate reconstruction shows a maximum in the last ~300 years during the Chilia III lobe construction, but storage rates may have increased earlier as the development of the St. George II lobe took place largely over the last millennium15 (Fig. 2A; SI). Late Holocene increases in delta plain expansion or sediment storage attributed to changes in land use have been similarly proposed for other European deltas such as the Po18,19,20 and the Rhone21.

Evolution of Danube-Black Sea system over the last 7,600 years (see SI for details on methodology and data).
Average storage rates for successive evolution phases of the delta (A) are compared to paleoevironmental proxies measured in core GGC18/MC19: alkenone δD composition as paleo-salinity proxies with salinity levels of ~30 and 20 psu calibrated for C-37 alkenones21 (B); 18S rDNA (as % of total reads) for diatoms (C) and dinoflagellates (D), principal phytoplankton taxa in the Black Sea, as indicators of food web structure. Lithostratigraphy is indicated by classical names of Black Sea units; black triangles show position of available radiocarbon dates (SI). The Holocene record of nitrogen fixation36 suggests increased continental input of N when δ15N is high (E). Holocene records of charcoal (F) and arboreal pollen (G) from Romanian low- and uplands32 track land use changes. Modeled deforestation rate that accounts for population history and technological advances40,41 is shown as percent of agricultural land in the Danube watershed in the lower basin (eastern Europe) and upper and middle basin (central Europe) (H).
Were the flux changes that led to the accelerated expansion of Danube delta in the late Holocene significant enough to affect the entire Black Sea? To address this question we used relevant proxies of past physical and biogeochemical conditions in the surface mixed layer (i.e., sea surface water salinity and abundance of major phytoplankton taxa) over the last ~9000 years (SI). Samples for analyses were collected in the western Black Sea (42°46.569″N; 28°40.647″E; 971 m water depth) on the continental slope south of the Danube mouths22. The Holocene sequence shows a typical Black Sea stratigraphy with oxic sediments at the base that were deposited in the low salinity “Black Sea” lake (Unit III), followed by an anoxic sapropel deposited after the ocean reconnection beginning at ~7,550 yrs BP (Unit II) (Fig. 2). At the top, Unit I consists of a laminated coccolith ooze deposited after ~2,560 yrs BP under conditions favorable for blooms of coccolith-bearing Emiliania huxleyi (Eh). This abrupt transition from sapropel to coccolith ooze suggests that an environmental threshold must have been crossed in the late Holocene23.
Previous paleogenetic studies at our site suggested that the occurrence of coccoliths22 as well as diversification in Eh viral population24 during the late Holocene may have been connected to changes in the hydrologic and nutrient regime of the Black Sea. Sea surface temperature, previously reconstructed on long-chain C37 alkenones22, shows a gradual cooling from ~19°C to 15°C from ~5000 toward 500 yrs BP. The δD composition of the same components (SI), measured for the present study (Fig. 2B), indicates a gradual increase in sea surface salinity in Unit IIa after ~5000 yrs BP toward a maximum of >30 psu at ~3000 yrs BP, followed by a decrease toward modern salinities close to 20 psu that characterized the last millennium (Fig. 2B). We also measured Eh-specific C36 alkenone δD composition (SI), which reinforces this sea surface salinity history and also suggests values fresher than ~20 psu between the base of Unit II and 5250 yrs BP where C37 alkenones are not reliable due to a variable haptophyte assemblage22.
An independent reconstruction in the eastern Black Sea23 also found a late Holocene decrease in surface waters salinity providing support for a general freshening of the basin during this time. Our modeled salinity history of the Black Sea (see SI) suggests that a higher fluvial influx alone would require an unrealistic increase by a factor of three to account for the late Holocene freshening. However, although direct precipitation over the Black Sea probably decreased in the late Holocene25, our model suggests that reduced evaporation during the gradual cooling over the same time period22 may have also contributed significantly to the freshening (SI). In addition, an augmented freshwater budget of the Black Sea due to increased fluvial discharge and reduced evaporation could have reinforced the freshening via hydrodynamic feedbacks. For example, the Mediterranean inflow through the Bosporus Strait is restricted when the freshwater balance is positive in the Black Sea13. Similarly, the mixed layer freshening inhibits mixing across the pycnocline12 limiting the exchange of salt with the deep waters of the Black Sea. Although we cannot quantify their contributions, an increase in fluvial discharge combined with decreased evaporation appears to be the probable cause for the late Holocene freshening of the Black Sea.
Precipitation reconstructions in the Black Sea drainage basin exhibit a considerable degree of regional variability as well as large century- to millennial-scale fluctuations25,26, which are not evident in the gradual decrease in Black Sea's salinity. The modeled increase in Danube's discharge16 between the coldest and wettest phase of the late Holocene – the Little Ice Age (LIA) – and the warmest and driest interval corresponding to latter half of 20th Century amounts to only ~10% of the average discharge. Similarly, modeled sediment fluxes for the Danube16 are also just marginally lower (~5%) during the LIA compared to the second half of 20th Century. This apparent lack of sensitivity implies that the transition from the warm and wet mid Holocene to the cold and dry late Holocene cannot explain any significant increases in water and sediment delivered to the Black Sea. Intensification of colluvial27, fluvial27,28, deltaic19,20,21 and shelf sedimentation18 in different climate settings in Europe points instead to land cover change as the main driver of continental scale sediment fluxes. However, increased climate variability in the late Holocene probably contributed to this intense denudation as the erosion sensitivity of landscapes may have increased due to land use changes29. Modern observations in large river basins7,30 also support the idea that a contemporaneous increase in freshwater delivery to the Black Sea probably accompanied the increase in sediment discharge, both caused by the massive human-controlled changes in land use during the late Holocene.
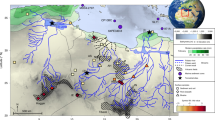
Historical records31 and land cover changes reconstructed using a scenario that accounts for both population history as well as technological advances suggest that the total deforested area in the lower Danube basin rapidly doubled after ~1,000 AD (Fig. 2H & 3). The land in use at the beginning of this rise was around 30–35% (SI). In contrast, Danube's upper and middle course were already deforested at a comparable level at the peak of the Roman Empire, 2000 years ago (Fig. 2H; SI). On the territory of modern Romania, which contributes close to 60% of the total Danube sediment load16, pollen studies suggest that tree cover diminished gradually after 3000 years ago in the lowlands (<300 m altitude), but more abruptly in the uplands in the last 1000 years32 (Fig. 2G). Biomass burning increased rapidly in the latter period, especially at altitudes higher than 800 m, but also in the lowlands32, supporting historical accounts of pasture and agricultural land expansion31 (Fig. 2F).

Modeled land use changes in the watershed of Danube River over the last millennium (white line separates upper and middle basin from the lower basin; see SI for details on modeling).
While the upper and middle basin was already heavily deforested at ~1000 A.D., rapid deforestation occurred in the lower basin after 1500 A.D. both prior as well as after the advent of the Industrial Revolution in the region after ~1850 A.D.
Changes in Danube's freshwater and sediment discharges should be reflected by changes in Black Sea's phytoplankton assemblages as nutrient fluxes may have also changed following anthropogenic disturbances such as forest clearance and agriculture2,33,34. Exceptional preservation of organic matter in the Black Sea under anoxic deposition with no bioturbation allows us to explore these changes by identifying and quantifying past plankton taxa based on their fossil sedimentary DNA (Fig. 2 and SI). We find that major phytoplankton taxa in the Black Sea changed drastically over the last millennium. The rise in diatoms after ~500 yrs BP suggests an increase of dissolved silicate input into the Black Sea surface waters (Fig. 2C). Consistent with an increase in stratification, the chemocline was shallower during the late Holocene freshening35. Based on modern circulation12, we assume that the nutrients are well mixed in the surface layer at scales longer then multiannual but we cannot exclude Si contributions from deep mixing during winter storms especially when the chemocline was shallow. However, such deep nutrient contributions documented to occur during the past 3000 years35 must have been minimal because they did not cause a concurrent rise in diatoms before 500 yrs BP. Instead, the ecological change in the last 500 years likely reflects an increased fluvial influx of dissolved silicate34 as a result of rapid deforestation in the lower Danube basin (Fig. 2 and 3; ref. 32 and SI). Dinoflagellate DNA also rose drastically after ~650 years ago (Fig. 2D), which is consistent with increased nutrient availability following anthropogenic disturbances such as forest clearance and agriculture2,33. Low nitrogen fixation in the Black Sea36 during that period supports an augmented fluvial N influx that coincides within chronological uncertainties with the rise in dinoflagellates (Fig. 2E). The earlier dinoflagellate DNA abundance peak during deposition of Unit IIb (~7000 to 5000 years ago) probably reflects increased nutrient contributions from below the chemocline35,36, although changes in phytoplankton communities linked to the gradual salinization of the basin or changes in the nutrient load of rivers draining into the Black Sea over that interval cannot be excluded. In contrast to the marine ecological changes following the rapid and extensive land use changes in the lower basin over that last ~500 years, the lack of an ecological response in the Black Sea to earlier deforestation in the upper and middle Danube basin points to possible onland buffering33 of distant and protracted land cover changes.
Discussion
Our combined sedimentary, paleoenvironmental and paleogenetic evidence suggests that land use changes transformed the Danube delta and the Black Sea well before the advent of the Industrial Revolution. Reconsidering the long term natural vs. human-induced influences should thus provide unique perspectives for management and conservation of coastal and marine resources. For the Black Sea, it remains to be assessed how changes detected in the ecosystem structure have affected biodiversity, productivity and biogeochemical cycling, but an unexpected positive legacy of this human impact history is the Danube delta itself. The foundation of its exceptional biodiversity37 is the great variety of environments including freshwater lakes, marshes, dunes, brackish lagoons and estuarine settings that were constructed in large part during the rapid delta growth phase in the late Holocene. At the global scale, coastal settings28,38 such as deltas, estuaries and marshes may have been similarly, if diachronously, altered by increased denudation3,9. Although the effects of ancient land use were probably more severe in marginal seas than in open ocean settings, the global role of these seas as large scale filters for material fluxes to the ocean may have been significant and remains to be assessed10.
Methods
Volumes of sediment accumulated in the Danube delta over the last ~9000 years and successive delta evolution phases15 were computed between the upper surface consisting of the modern delta plain topography and bathymetry down to −40 m water depth and a lower surface defined by the pre-Holocene delta plain39. Mass estimates use a dry density of 1.5 g cm−3 for water saturated mixed sand and mud with 40% porosity (see SI for details). We extracted historical estimates of land use in the Danube basin from the KK10 standard scenario of Holocene anthropogenic land cover change17,40,41 (SI). Changes in Black Sea salinity were modeled using a simplified time-stepping simulation of the two-way flow through the Bosphorus42 (see SI). The Manning equation for open-channel flow was used to describe the outflow and the Poiseuille equation for pipe flow was adopted for the inflow.
Sedimentary alkenone δD, a proxy for salinity in the Black Sea23, was measured using an Agilent 6890 GC equipped with a DB-5MS column (60 m length; 250 µm i.d.; 0.25 µm film thickness; J&W, Folsom, CA, USA) coupled to a DeltaPlus XL (Bremen, Germany) isotope ratio mass spectrometer via a pyrolysis interface operated at 1440°C (see SI). Total genomic DNA from 44 selected sediment intervals used in other (paleo)genetic studies (e.g., reference 24), served as template for PCR amplification of environmental eukaryotic 18S rRNA gene fragments (V1 to V3 region) and subsequent 454 GS FLX Titanium pyrosequencing using facilities of Engencore (Columbia, SC) and conditions as described in detail in the SI. Each 1/8 section of the 454 picotiterplate contained 15 pooled barcoded PCR products. Dereplication and taxonomic analysis of remaining high quality sequence reads (500–6000 sequence reads per sample, 360 bp in length) was done using the PyroTagger pipeline (http://pyrotagger.jgi-psf.org/).
References
Turner, R. E. & Rabalais, N. N. Coastal eutrophication near the Mississippi river delta. Nature 368, 619–621 (1994).
Humborg, C., Ittekkot, V., Cociasu, A. & VonBodungen, B. Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature 386, 385–388 (1997).
Syvitski, J. P. M., Vörösmarty, C. J., Kettner, A. J. & Green, P. Impact of humans on the flux of terrestrial sediment to the global coastal ocean. Science 308, 376–380 (2005).
Zalasiewicz, J., Williams, M., Steffen, W. & Crutzen, P. The new world of the Anthropocene. Env. Sci. & Tech. 44, 2228–2231 (2010).
Ruddiman, W. F. The anthropogenic greenhouse era began thousands of years ago. Climatic Change 61, 261–293 (2003).
Doney, S. C. The growing human footprint on coastal and open-ocean biogeochemistry. Science 328, 1512–1516 (2011).
Raymond, P. A., Oh, N., Turner, E. R. & Broussard, W. Anthropogenically enhanced fluxes of water and carbon from the Mississippi River. Nature 451, 449–452 (2008).
Zillen, L. & Conley, D. J. Hypoxia and cyanobacteria blooms - are they really natural features of the late Holocene history of the Baltic Sea? Biogeosciences 7, 2567–2580 (2010).
Wilkinson, B. H. & McElroy, B. J. The impact of humans on continental erosion and sedimentation. GSA Bull. 119, 140–156 (2007).
Meybeck, M., Dürr, H. H., Roussennac, S. & Ludwig, W. Regional seas and their interception of riverine fluxes to oceans. Mar. Chem. 106, 301–325 (2007).
Kideys, A. E. Fall and rise of the Black Sea ecosystem. Science 297, 1482–1484 (2002).
Özsoy, E. & Ünlüata, Ü. Oceanography of the Black Sea: a review of some recent results. Earth Sci. Rev. 42, 231–272 (1997).
Stanev, E. V. & Peneva, E. L. Regional sea level response to global climatic change: Black Sea examples. Global Planet. Change 32, 33–47 (2001).
Degens, E. T., Kempe, S. & Richey, J. E. Biogeochemistry of Major World Rivers (John Wiley and Sons, 1991).
Giosan, L. et al. Young Danube delta documents stable Black Sea level since the middle Holocene: morphodynamic, paleogeographic and archaeological implications. Geology 34, 757–760 (2006).
McCarney-Castle, K., Voulgaris, G., Kettner, A. J. & Giosan, L. Simulating fluvial fluxes in the Danube watershed: The Little Ice Age versus modern day. Holocene 22, 91–105 (2012).
Kaplan, J. O. et al. Holocene carbon emissions as a result of anthropogenic land cover change. Holocene 21, 775–791 (2011).
Oldfield, F. et al. A high resolution late Holocene palaeo environmental record from the central Adriatic Sea. Quat. Sci. Rev. 22, 319–342 (2003).
Correggiari, A., Cattaneo, A. & Trincardi, F. The modern Po Delta system: Lobe switching and asymmetric prodelta growth. Mar. Geol. 222–223, 49–74 (2005).
Brommer, M. B., Weltje, G.-J. & Trincardi, F. Reconstruction of sediment supply from mass accumulation rates in the Northern Adriatic Basin (Italy) over the past 19,000 years. J. Geophys. Res. 114, F02008 (2009).
Vella, C. et al. Evolution of the Rhone delta plain in the Holocene. Mar. Geol. 222–223, 235–265 (2005).
Coolen, M. J. L. et al. DNA and lipid molecular stratigraphic records of haptophyte succession in the Black Sea during the Holocene. Earth Planet. Sci. Lett. 284, 610–621 (2009).
Van der Meer, M. T. J. et al. Molecular isotopic and dinoflagellate evidence for Late Holocene freshening of the Black Sea. Earth Planet. Sci. Lett. 267, 426–434 (2008).
Coolen, M. J. L. 7000 Years of Emiliania huxleyi Viruses in the Black Sea. Science 333, 451–452 (2011).
Goektuerk, O. M. et al. Climate on the southern Black Sea coast during the Holocene: implications from the Sofular Cave record. Quat. Sci. Rev. 30, 2433–2445 (2011).
Buentgen, U. et al. 2500 years of European climate variability and human susceptibility. Science 331, 578–582 (2011).
Dotterweich, M. The history of soil erosion and fluvial deposits in small catchments of central Europe: Deciphering the long-term interaction between humans and the environment — A review. Geomorphology, 101, 192–208 (2008).
Hoffmann, T., Erkens, G., Gerlach, R., Klostermann, J. & Lang, A. Trends and controls of Holocene floodplain sedimentation in the Rhine catchment. Catena 77, 96–106 (2009).
Lang, A. et al. Changes in sediment flux and storage within a fluvial system: some examples from the Rhine catchment. Hydrol. Process. 17, 3321–3334 (2003).
Coe, M., Costa, M. & Soares-Filho, B. S. The influence of historical and potential future deforestation on the stream flow of the Amazon River—land surface processes and atmospheric feedbacks. J. Hydrology 369, 165–174 (2009).
Giurescu, C. C. Istoria Pădurii Româneşti (Editura Ceres, 1976).
Feurdean, A. et al. Trends in biomass burning in the Carpathian region over the last 15,000 years. Quat. Sci. Rev. 45, 111–125 (2012).
McLauchlan et al. Changes in nitrogen cycling during the past century in a northern hardwood forest. PNAS 104, 7466–7470 (2007).
Struyf et al. Historical land use change has lowered terrestrial silica mobilization. Nature Comm. 1, 129 (2010).
Damste, J. S. S. et al. A 6,000-year sedimentary molecular record of chemocline excursions in the Black Sea. Nature 362, 827–829 (1993).
Fulton, J. M., Arthur, M. A. & Freeman, K. H. Black Sea nitrogen cycling and the preservation of phytoplankton δ15N signals during the Holocene. Global Biogeochem. Cycles 26, GB2030 (2012).
Sommerwerk, N. et al. in Rivers of Europe. (eds Tockner, K., Robinson, C. & Uehlinger, U.) 59–112 (Academic Press., 2009).
Kirwan, M. L., Murray, A. B., Donnelly, J. P. & Corbett, D. R. Rapid wetland expansion during European settlement and its implication for marsh survival under modern sediment delivery rates. Geology 39, 507–510 (2011).
Giosan, L., Filip, F. & Constantinescu, S. Was the Black Sea catastrophically flooded in the early Holocene? Quat. Sci. Rev. 28, 1–6 (2009).
Kaplan, J. O., Krumhardt, K. M. & Zimmerman, N. E. The prehistoric and preindustrial deforestation of Europe. Quat. Sci. Rev. (2009).
Kaplan, J. O., Krumhardt, K. M. & Zimmerman, N. E. The effects of human underland use and climate change over the past 500 years on the carbon cycle of Europe, Glob. Change Biol. 10.1111/j.1365–2486.2011.02580.x (2011).
Thom, N. A hydrologic model of the Black and Caspian Seas in the late Pleistocene and early-middle Holocene. Quat. Sci. Rev. 29, 2989–2995 (2010).
Acknowledgements
We thank colleagues from University of Bucharest and the Danube Delta Institute in Romania for support in the field in the Danube delta and colleagues from IOBAS Varna in Bulgaria for support during the cruise aboard R/V Akademik. We thank Sean Sylva at WHOI for technical expertise. This study was supported by grants OISE 0637108, EAR 0952146, OCE 0602423 and OCE 0825020 from the National Science Foundation and grants from the Woods Hole Oceanographic Institution.
Author information
Authors and Affiliations
Contributions
The project was initiated by L.G. and M.C. and designed by L.G., M.C., J.O.C. and A.K. J.O.C. and N.T. contributed the land use and hydrological modeling components respectively. Field and lab data were collected by L.G., M.C., S.C., F.F. and M.F.-M. Danube delta data were analyzed and interpreted by S.C., L.G. and F.F. L.G. wrote the paper with contributions from all authors.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Giosan, L., Coolen, M., Kaplan, J. et al. Early Anthropogenic Transformation of the Danube-Black Sea System. Sci Rep 2, 582 (2012). https://doi.org/10.1038/srep00582
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00582
This article is cited by
-
World’s largest dam removal reverses coastal erosion
Scientific Reports (2019)
-
Impacts of forest loss in the eastern Carpathian Mountains: linking remote sensing and sediment changes in a mid-altitude catchment (Red Lake, Romania)
Regional Environmental Change (2019)
-
Historical ecology of riverine fish in Europe
Aquatic Sciences (2015)
-
Man made deltas
Scientific Reports (2013)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
