
Abstract
We describe a previously unknown assemblage of seamount-associated megabenthos that has by far the highest peak biomass reported in the deep-sea outside of vent communities. The assemblage was found at depths of 2–2.5 km on rocky geomorphic features off the southeast coast of Australia, in an area near the Sub-Antarctic Zone characterised by high rates of surface productivity and carbon export to the deep-ocean. These conditions and the taxa in the assemblage, are widely distributed around the Southern mid-latitudes, suggesting the high-biomass assemblage is also likely to be widespread. The role of this assemblage in regional ecosystem and carbon dynamics and its sensitivities to anthropogenic impacts are unknown. The discovery highlights the lack of information on deep-sea biota worldwide and the potential for unanticipated impacts of deep-sea exploitation.
Similar content being viewed by others

Introduction
Conventional wisdom and extensive data from deep-sea dredging worldwide indicate that biomass densities in the deep ocean are generally low, often peaking at or around the shelf edge and then declining with increasing depth1,2. Noteworthy exceptions are dense pockets of chemosynthetic organisms associated with hydrothermal vents3,4, scavengers aggregated around ephemeral food sources, such as whale carcasses5 and infauna at the top of shelf canyon features, apparently supported by topographic focussing of coastal carbon sources transported offshore6. In December 2008/January 2009, we discovered a new, exceptionally high biomass marine community while undertaking a series of dives by the Remotely Operated Vehicle Jason on seamounts and adjacent geomorphic features off southern Tasmania, Australia (Figure 1). This paper describes the new assemblage, comments on the production sources likely to be supporting it and speculates on the possible roles of the assemblage in regional ecosystem dynamics.

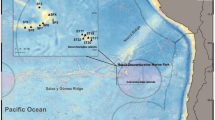
Map of study area, showing towed camera, Jason and ABE dive locations off SE Australia.
The sample sites are roughly bounded by 44–46° S and 144–147° E.
Results
The Tasmanian seamounts are of volcanic origin, come to within about 750 m of the surface and support an extensive benthic community, dominated at depths < 1300 m by the reef-forming stony corals (scleractinians), Solenosmilia variabilis and Enallopsammia rostrata7,8. Initial, relatively shallow dives in the seamount area appeared to confirm previous suggestions that biomass distributions were highest near the seamount peaks and declined with increasing depth, with biota becoming increasingly sparse to approximately 1800 m7. However, below this depth the abundance of benthic organisms abruptly increased, as a result of the presence often in huge numbers of three taxonomically unrelated benthic organisms: large barnacles (apparently Tetrachaelasma tasmanicum9), an undescribed hormathiid anemone and large bamboo corals (gorgonians) in the isidid genera Keratoisis and Isidella, also undescribed (Figure 2). Neither the barnacle nor anemone had previously been seen or collected in shallower surveys. Thousands of both were often visible from the Jason cameras, covering most available rock surfaces. The organisms were conspicuously alive, verified by numerous specimens collected using the Jason manipulator arms. Reflecting their evident abundance and contrary to expectations, the number of visible megabenthic organisms seen in the Jason images averaged almost 3 times higher between 2000 and 2500 m than in comparable images for the scleractinian reef-assemblage between 1000 and 1300 m (116.8 vs. 29.3 individuals/image, based on 117 and 1059 images, respectively)(Fig. 3).

(a) and (b).
Hormathiid-dominated assemblage on volcanic rock, at 2400 m and 2360 m, respectively. Jason dive #392, at 45° 17.75′′ S; 146° 06.33′′ E. The rubble surrounding the rock is predominantly barnacle skeletal plates. Live barnacles are visible as white objects amongst the greenish-brown anemones; to the left are two branching isidid colonies. For scale, the anemones are about 5 cm in diameter. (c). Isidid gorgonians (Keratoisis spp) and anemones at 2540 m. Jason dive #392, at 45° 17.70′′ S; 146° 06.72′′ E.

Megabenthos abundance as a function of depth, binned by 100 m depths, based on counts made from the Jason “light-bar camera” and comparable still images from the CSIRO towed camera system8.
Mean area of the images was 5.1 m2. Thin horizontal lines indicate the minimum and maximum, the thick line the standard deviation and the thin vertical line the mean for each depth range.
Biomass in the deep zone varied widely, in large part depending on the proportion of rocky substrata in each image, but was often extremely high. We estimate a peak value for the assemblage of 16.9 kg/m2 shell-free wet weight, based on an image at 2056 m that contained 619 anemones, 30 gorgonian colonies and 27 barnacles and average shell-free wet weights of preserved specimens of 24.1, 42.9 and 19.8 g, respectively (see Methods). By comparison, maximum whole organism wet weight density for the shallower scleractinian reef was recently reported to be about 2.4 kg/m2 10, from which we estimate a peak shell-free biomass of about 0.4 kg/m2. The highest biomass density we observed on the shallow reef was 2.1 kg/m2 shell-free wet weight, based on 528 Enallopsammia rostrata colonies in an image at 866 m.
The source of the production supporting this high biomass assemblage is not known. For a preliminary indication of this source, we measured ∂14C in freeze-dried wet tissue collected from live-caught specimens of all three of the abundant deep taxa and compared them with regional ∂14C profiles of dissolved inorganic carbon (DIC) and suspended particulate organic carbon (POC) (Figure 4). The radiocarbon analyses show post-bomb carbon in all three taxa and ∂14C values in the lower end of the range of surface DIC. These values are consistent with a diet based on sinking particulates derived from recent surface production11,12 and either suspended POC or adsorption of low levels of DOC on to sinking particulates13,14.

∂14C values of freeze-dried wet tissue obtained from live specimens of isidid gorgonians (G), barnacles (B) and hormathiid anemones (A), collected using the Jason and frozen at −70C° prior to analysis, compared to water column profiles of dissolved inorganic carbon (DIC) in the Tasman Sea (latitude band 43–46° S) just east of the seamount region, from ref. 26 and the range of ∂14C values reported by ref. 27 for particulate organic carbon (POC)(particles > 1 µm diameter) in the Southern Ocean southeast of the study area (54° S, 176° W).
Discussion
The density and biomass of organisms we found at 2000–2500 m is an order of magnitude higher than any previous reports in this depth range and dwarfs even the exceptional values recently reported for in-fauna at the mouths of submarine canyons off New Zealand (maximum wet weight 2.1 kg/m2)6. Wet weight biomass density is similar to and even exceeds some reports for vent communities3,4, but covers a much larger surface area and hence represents a substantially higher overall biomass. Proximally, the high anemone density could be facilitated by asexual reproduction, which is common in the Order15. Little is known about the reproductive biology or aggregative behaviour of deep-sea barnacles and gorgonians. Ultimately, the high densities need to be supported by a significant production source. Radiocarbon analyses of freeze-dried wet tissues are consistent with a diet based wholly or primarily on recent surface production and sinking particulates. The source of this production is not known, but it is perhaps noteworthy that the seamounts are located close to the Sub-Antarctic Zone in the Tasman Sea, an area of high productivity and carbon export to the deep ocean16,17. The peak benthic biomass on the deep reef, approximately 1.2 kg/m2 C, is more than an order of magnitude higher than would be expected based on surface primary production regionally (300 – 600 g C/m2/year17) and the 1–2 trophic levels between the gorgonians, at least and surface production12. The implications are either a large, but unknown C source supporting the assemblage, a mechanism as yet unidentified that that focuses export production at 2 –2.5 km depth, or very low biomass turnover rates11,12.
The geographic extent of the deep high-biomass assemblage is not known due to very sparse benthic sampling on rocky substrata globally in this depth range. The assemblage was seen on dives throughout the sampled area, but the highest densities tended to be in the western half. The significance of this, if any, is not known. However, the taxa are not unique to the Tasmanian region, similar deep rocky substrata occur widely on seamount features in the Southern hemisphere18 and high rates of export of surface carbon to depths >2000 m occur around the hemisphere in this latitude band19. These observations suggest that the assemblage may be widespread around the temperate margin of the Southern Ocean.
The roles of this newly discovered deep-sea faunal assemblage in regional ecosystem dynamics and in sequestration of carbon exported to the deep ocean from the Sub-Antarctic Zone17,20 are unknown. If widespread, the huge biomasses involved in this assemblage could constitute a significant, previously unknown factor in sequestration of Southern Ocean surface production. Reconstruction of historical variability of at least one of the three components, the barnacle, may be possible from the very extensive deposits of its calcite skeletal plates at 2 – 2.5 km depths (Fig. 2). These deposits are well above the calcite saturation horizon (3.2 km)21 and hence could constitute a substantial long-term archive. Irrespective of its paleo-oceanographic and ecological significance, the presence of this previously unknown extraordinary assemblage highlights the potential for unforseen negative consequences of rapidly developing deep-sea exploitation in unexplored national and International Waters22,23. Deep-sea trawling and oil, gas and mineral exploration and extraction now take place at the depths occupied by the assemblage24. The impact of these activities on what are likely to be very slow-growing individuals is potentially profound and long lasting. Our observations suggest that an assumption that biota at these depths is sparse and hence that biological impacts of resource exploitation is likely to be slight is not warranted.
Methods
Sampling vehicle descriptions and further information about the Tasmanian seamount region, its geomorphology and biota can be found at www.cmar.csiro.au/research/seamounts/video and www.whoi.edu/page.do?pid=8421, respectively. Information from the Jason dives, which spanned a depth range of 729 to 4011 m, was supplemented by photographic surveys of the area to 2950 m in January 2008 using the Woods Hole Oceanographic Institution Autonomous Benthic Explorer (ABE) and to 1490 m in two sister surveys in November 2006/April 2007 using the CSIRO Deep Towed Camera System8. Biomass densities were estimated from quantitative planar images of approximately 16 m2 taken using ABE, as it maintained a 3 m altitude above the substratum.
Live specimens were collected at numerous sites using the Jason manipulator arms. The anemones were identified by D. Fautin (Univ. Kansas) and M. Mitchel (Museum of Victoria), the gorgonians by P. Alderslade (CSIRO) and the barnacle by K. Gowlett-Holmes (CSIRO) from preserved material.
Shell/skeleton-free wet weights were determined as follows
Hormathiids
Twelve specimens from 7 different sampling stations, preserved in 90% ethanol, were blotted dry and weighed to the nearest 0.1 g. Specimen weights ranged from 4.2 to 43.1 g, averaging 18.8 g. Ethanol is 77% as dense as seawater, suggesting a true mean wet weight in seawater of 24.1 g (see discussion in ref 25).
Bamboo coral
Live tissue (polyps and connecting tissue) was stripped from a 5.5 cm branch (internode) of an ethanol (90%) preserved specimen, of about the same branch diameter as those observed from the Jason, blotted dry and weighed to the nearest 0.01 g. Total wet weight of the tissue in ethanol was 1.85 g, which equates to 0.33 g/cm of skeleton. We estimated the average height of a bamboo coral colony at 2000–2500 m to be about 0.75 m and from photographs conservatively estimated the total linear branch length per colony to be about 2 m and the proportion covered by live tissue to be about 50%. The total ethanol preserved wet weight per colony, therefore, was 0.33 g/cm × 200 cm × 0.5 = 33 g. Adjusting for ethanol preservation suggests a wet weight in seawater of 42.9 g.
Barnacle
An average sized barnacle collected using Jason at 2217 m and frozen at −70° C weighed 60 g. This was converted to an estimated shell-free dry weight using the cirripede conversion ratio of 0.066% from ref. 25 and then scaled up to wet weight using an approximate figure from their paper of 0.2 dry weight/wet weight. The total estimated shell-free wet weight of the specimen, therefore, was 60 g × 0.066 × 5 = 19.8 g.
Solenosmilia variabilis and Enallopsammia rostrata
Three growing tips of S. variabilis that clearly contained polyps were cut from a colony live-collected using Jason at approximately 1000 m depth. The specimen had been frozen at −70° C immediately after collection. The tips were cut off using a Dremel saw and weighed to the nearest 0.001 g. Specimen weights were 1.917, 2.495 and 4.286 g. The specimens were thawed and decalcified in 0.5 M EDTA, pH adjusted to 8, until all shell material was dissolved. The remaining tissue was rinsed in deionised water, blotted and weighed. Ratios of shell-free wet weight to total wet weigh were 0.20, 0.094 and 0.103, respectively, for an average of 0.133. Rowden et al10 report a maximum total wet weight for the Tasmanian Solenosmilia reefs of 2.4 kg/m2. We apply the conversion ratio of 0.133 to their estimated 73% of the total assemblage wet weight made up of live S. variabilis, on a generous assumption that each colony is entirely filled with polyps and then increase the total shell-free biomass by 50% to account for the remaining diverse species. Most of these are cnidarians, which are likely to have an equal or lower conversion ratio than S. variabilis.
The estimate of wet weight biomass of E. rostrata was based on the weight of a whole, large colony (360 g) collected at 870 m using Jason and frozen immediately at −70 C. We assume a generous ratio of shell-free wet weight to total wet weigh of 0.20.
Peak biomass density in g C for the deep assemblage was calculated using ratios of AFDW/WW of 0.1525 and C/DW of 0.5.
Radiocarbon analyses follow procedures in Sherwood, et al12.
References
Rex, M. A., et al. Global bathymetric patterns of standing stock and body size in the deep-sea benthos. Mar. Ecol. Prog. Ser. 317, 1–8 (2006).
Lampitt, R. S., Billett, D. S. M., Rice, A. L. Biomass of the invertebrate megabenthos from 500 to 4100 m in the northeast Atlantic Ocean. Mar. Biol. 93, 69–81 (1986).
Biscoito, M., Cary, C., Dixon, D. & Wilson, C. Proceedings of the First International Symposium on Deep-sea Hydrothermal Vent Biology. Cah. Biol. Mar. 39, 219–392 (1998).
Lutz, R. A. & Kennish, M. J. Ecology of deep-sea hydrothermal vent communities: a review. Rev. Geophys. 31, 211–241 (1993).
Haedich, R. L. & Rowe, G. T. Megafaunal biomass in the deep sea. Nature 269, 141–142 (1977).
De Leo, F. C., Smith, C. R., Rowden, A. A., Bowden, D. A. & Clark, M. R. Submarine canyons: hostpots of benthic biomass and productivity in the deep sea. Proc. Royal Soc. (Lond). B, Biol. Sci. 277, 2783–2792 (2010).
Koslow, J. A., et al. Seamount benthic macrofauna off southern Tasmania: community structure and impacts of trawling. Mar. Ecol. Prog. Ser. 213, 111–125 (2001).
Althaus, F., et al. Impacts of bottom trawling on deep-coral ecosystems of seamounts are long-lasting. Mar. Ecol. Prog. Ser. 397, 279–294 (2009).
Buckeridge, J. S. A new deep sea barnacle, Tetrachaelasma tasmanicum sp. nov. (Cirripedia: Balanomorpha) from the South Tasman Rise, South Pacific Ocean. New Zealand J. Mar. Fresh. Res. 33, 521–531 (1999).
Rowden, A. A., et al. A test of the seamount oasis hypothesis: seamounts support higher epibenthic megafaunal biomass than adjacent slopes. Mar. Ecol. 31 (Supp. s1), 95–106 (2010).
Roark, E. B., et al. Radiocarbon-based ages and growth rates of bamboo corals from the Gulf of Alaska. Geophys. Res. Lett. 32, L04606. 10.1029/2004GL021919 (2005).
Sherwood, O. A., Thresher, R. E., Fallon, S. J., Davies, D. M. & Trull, T. W. Tasmanian isidids: isotopic evidence for benthic-pelagic coupling and centennial constancy of oceanographic conditions. Mar. Ecol. Prog. Ser. 397, 209–218 (2009).
Druffel, E. R. M. & Williams, P. M. Identification of a deep marine source of particulate organic carbon using bomb 14C. Nature 347, 172–174 (1990).
Hwang, J., Druffel, E. R. M. & Bauer, J. E. Incorporation of aged dissolved organic carbon (DOC) by oceanic particulate organic carbon (POC): an experimental approach using natural carbon isotopes. Mar. Chem. 98, 315–322 (2006).
Fautin, D. G. Reproduction in Cnidaria. Can. J. Zool. 80, 1735–1754 (2002).
Clementson, L. A., et al. Controls on phytoplankton production in the Australasian sector of the subtropical convergence. Deep-sea Res. I 45, 1627–1661 (1998).
Mongin, M., Matear, R. & Chamberlain, M. Simulation of chlorophyll and iron supplies in the Sub Antarctic Zone South of Australia. Deep-sea Res. I (press 01).
Kitchingman, A., Lai, S., Morato, T. & Pauly, P. How many seamounts are there and where are they located. Pp. 26–40 In: Pitcher T. J., et al. (Seamounts:Ecology, Fisheries & Conservation. Blackwell, Oxgford, UK. (2007).
Lutz, M. J., Caldeira, K., Dunbar, R. B., & Behrenfield, M. J. Seasonal rhythms of net primary production and particulate organic carbon flux describe biological pump efficiency in the global ocean. J. Geophys. Res. 112, C10011 (2007).
Lourey, M. J., Trull, T. W. & Sigman, D. M. Sensitivity of ∂15N of nitrate, surface suspended and deep sinking particulate nitrogen to seasonal nitrate depletion in the Southern Ocean. Global Biogeochem. Cycles 17, 1–18 (2003).
Thresher, R. E., Thresher, R. E., Tilbrook, B., Fallon, S., Wilson, N. C. & Adkins, J. Effects of chronic low carbonate saturation levels on the distribution, growth and skeletal chemistry of deep-sea corals and other seamount megabenthos. Mar Ecol. Prog. Ser. (press 01).
Glover, A. G. & Smith, C. R. The deep-sea floor ecosystem: current status and prospects of anthropogenic change by the year 2025. Env. Cons. 30, 219–241 (2003).
Hein, J. R., Conrad, T. A. & Staudigel, H. Seamount mineral deposits: a source of rare metals for high-technology industries. Oceanography 23, 184 – 189 (2010).
Ramirez-Llodra, E., et al. Man and the last great wilderness:human impact on the deep sea. PLOS One 6(7): 25 Pp (2011).
Ricciardi, A. & Bourget, E. Weight-to-weight conversion factors for marine benthic macroinvertebrates. Mar. Ecol. Prog. Ser. 163, 245–251 (1998).
Lassey, K. R., Manning, M. R., Sparks, R. J. & Wallace, G. Radiocarbon in the Sub-tropical Convergence east of Tasmania- an interim report. Dept. Science and Industrial Research (New Zealand) Physical Sciences Report 11: 14 Pp (1990).
Druffel, E. R. M. & Bauer, J. E. Radiocarbon distributions in Southern Ocean dissolved and particulate organic matter. Geophys. Res. Lett. 27, 1495–1498 (2000).
Acknowledgements
We thank E. Anagnostou, B. Barker, A. Beck, W. Cho, J. Cordell, A. Gagnon, R. Gurney, S. John, A. Kennedy, H. Kippo, M. Lewis, F. McEnnulty, N. Meckler, D. Mills, M. Sherlock, D. Staples, A. Subhas and N. Thiagarajan and the crews of the AUV ABE, the ROV Jason and RV Thomas T. Thompson and RV Southern Surveyor for their invaluable and professional assistance in the field, P. Alderslade, S. Cairns, D. Fautin, M. Mitchel and K. Moore for taxonomic assistance, P. Brodie and J. Dowdney for assisting in the photographic analyses, T. Trull for discussion regarding the radiocarbon analyses, J. Guinotte and R. Matear for discussions on regional patterns of productivity and export and N. Bax, F. McEnnulty and the NSR sub-editor, M. Scardi, for valuable comments on the ms. This work was supported by the U.S. National Science Foundation, the Australian Department of Environment, Water, Heritage and the Arts, the Australian Commonwealth Environmental Research Fund Marine Biodiversity Hub and a grant of ship time by the Australian National Research Facility.
Author information
Authors and Affiliations
Contributions
R.E.T. and J.A. organised, coordinated and secured support for the Jason and ABE field programs and with K. G.-H. undertook the field observations; S.J.F. did the radiocarbon analyses; F.A. and A.W. oversaw the quantitative analysis of photographic images; and R.E.T. did the wet weight analyses. All five authors contributed to writing the ms.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Thresher, R., Adkins, J., Fallon, S. et al. Extraordinarily high biomass benthic community on Southern Ocean seamounts. Sci Rep 1, 119 (2011). https://doi.org/10.1038/srep00119
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00119
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
