


















« Prev Next »

Succession refers to a directional, predictable change in community structure over time (Grime 1979, Huston & Smith 1987). This change is due to shifts in the presence and relative abundance of different species as time passes over years to centuries. While succession is most often thought about in terms of the plant community, shifts in the populations of other organisms also need to be considered. The process of succession can be seen in many different systems, ranging from the establishment of grasslands after a volcanic eruption, to the re-establishment of forests after agricultural fields have been abandoned.
The Role of Disturbance
In plant communities, succession begins when an area is made partially or completely devoid of vegetation because of a disturbance. Disturbances can take many different forms, and can vary in intensity and size. Large, extreme disturbances such as volcanic eruptions or glacier retreat result in very slow succession due to complete mortality of all living individuals in the system, as well as loss of the entire soil complex. This type of succession is often called primary succession, as the soil, as well as all characteristic organisms, needs to re-establish. Large, low intensity disturbances, such as plowing in conventional agriculture, result in moderate amounts of succession, where species can survive in the soil and quickly recolonize areas after a disturbance. This type of succession is often called secondary succession (Connell & Slayter 1977). Early successional communities are those that establish themselves relatively quickly after disturbance, while late successional communities are those that establish themselves much later.
Classic Perspectives of Succession
Succession is one of the longest-studied ecological concepts. Henry Cowles was the first ecologist to thoroughly characterize successional patterns, which he did in his classic 1899 study of sand dunes along the shores of Lake Michigan (Cowles 1899). Cowles described the chronosequence of vegetation along sand dunes, moving from bare sand beach, to grasslands, to mature forests. A chronosequence is a “space-for-time” substitution, where ecologists can predict temporal patterns of vegetation based on a snapshot of an area gradient representing different ages of succession (Figure 1).
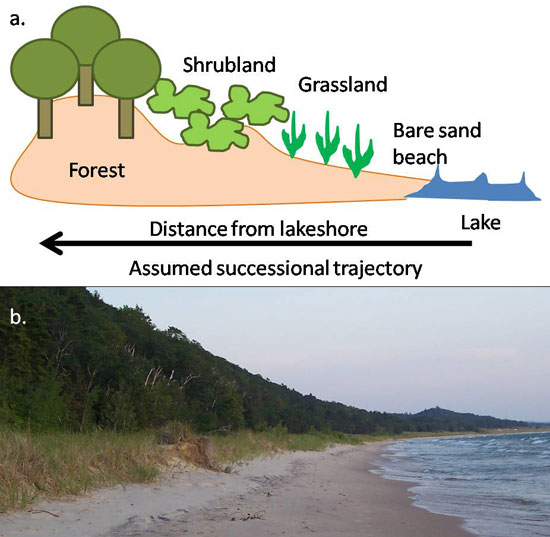
Figure 1: Chronosequences are often used to study succession
(A) Typical chronosequence for sand dune succession. (B) Sleeping Bear Dunes National Lakeshore, MI showing successional chronosequence from bare sand to grassland to forest.
The concept of predictable change in vegetation time was next championed by Frederick Clements in the early 1900s. He proposed the concept of a climax state for communities, which represented the final, or permanent, end-stage of succession (Clements 1936). For Clements, climax communities were the assemblage of characteristic plants that define an ecosystem, such as tall grasses in a prairie, or mature trees in a forest. Clements held that, after a disturbance, any given ecosystem would eventually return to its characteristic assemblage of species. For example, if an oak-hickory forest had a severe forest fire which destroyed most of the trees, that forest system would eventually return to the climax community, defined by oak- and hickory-dominated species. Clements’s ideas of the extreme predictability of succession led him to propose a super-organism concept of succession, whereby all species in the climax community work together to maintain a stable composition (Figure 2). This idea, that an ecosystem could self-form, or self-renew into a stable climax community, became very popular in the 1920s.

Figure 2: Two contrasting views of succession
(A) The super-organism concept, where groups of species are tightly associated, and are supplanted by other groups of tightly associated species. (B) The individualistic concept, where individual species independently respond to environmental conditions. Each curve on the graphs represents the abundance of a single species.
While the concept of a climax community is still viable today, the super-organism concept was opposed by another ecologist, Henry Gleason. Gleason argued that communities were individualistic; that is, communities were only the fortuitous assembly of species, and that there was no such thing as a climax state for ecosystems. Gleason recognized that the environment, and species’ movements, had an important role in regulating species assemblages, and that community changes were not nearly as predictable as Clements had proposed (Gleason 1926). While Gleason’s ideas were not well received by scientists in the 1920s, his recognition of the random aspects of community assembly are appreciated today by community ecologists interested in neutral models of biodiversity (e.g., Hubbell 2001).
Patterns and Mechanisms of Succession

Figure 3: Changes over time in total plant species richness over time at select sites on Mount Saint Helens, WA
Plant reestablishment 15 years after the debris avalanche at Mount St. Helens, Washington. (Figure modified from Dale & Adams 2002)
It wasn’t until 1977 that ecologists actually proposed mechanisms by which communities might progress through predictable successional sequences. Facilitation is the most common mechanism proposed to explain succession. This occurs when one species, or a group of species, colonizes a disturbed area, and subsequently alters the environment of that area (by altering soil nutrients, light accessibility, or water availability), making it more habitable for later successional species (Connell & Slayter 1977). However, other possible mechanisms included tolerance, inhibition, and random colonization.
One of the best examples of primary succession comes from studies by William Cooper, William Reiners, Terry Chapin, and others in Glacier Bay, Alaska (Cooper 1923, Reiners et al. 1971, Chapin et al. 1994). Since 1794, the glacier filling Glacier Bay has steadily been retreating (Figure 4a). Researchers have characterized primary succession in this system, where plant communities progress from pioneer species (i.e., early colonizing lichens, liverworts, and forbs) to creeping shrubs such as Dryas, to larger shrubs and trees such as alder, and finally to the climax spruce forest community over 1,500 years (e.g., see Chapin et al. 1994). Both facilitation and inhibition act as mechanisms regulating succession in this system (Figure 4b). For example, both Dryas and alders increase soil nitrogen, which increases the establishment and growth of spruce seedlings. However, both Dryas and alders produce leaf litter which can inhibit spruce germination and survival.
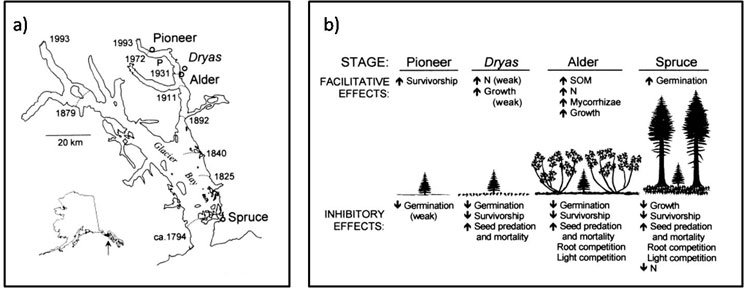
Figure 4: Succession after glacier retreat
(A) Map of glacier retreat from 1794-1993 in Glacier Bay, AK. (B) Summary of facilitative and inhibitory effects of each successional stage of vegetation on spruce seedling growth. (Figure modified from Chapin et al. 1994)
A classic study of secondary succession was conducted by Catherine Keever (1950). In this study, Keever characterized succession in an old field after agricultural use had ceased. She observed a predictable shift in plant community composition following field abandonment, with horseweed (Erigeron canadense) dominating fields one year after abandonment, white aster (Aster pilosis) dominating in year two, and broomsedge (Andropogon virginicus) dominating in year three (Figure 5). She found that life history strategies of individual species, seed dispersal, allelopathy (biochemical production by a plant which alters growth and survival of other plants or itself), and competitive interactions among species, led to this predictable pattern of succession. Both Chapin’s and Keever’s studies clearly demonstrate that multiple mechanisms can operate during the process of succession.
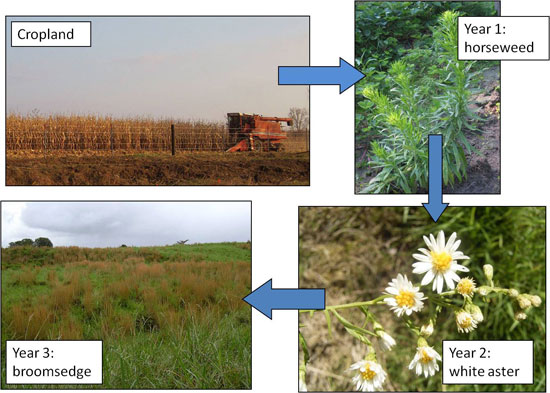
Figure 5: Keever's observed pattern of succession in North Carolina agricultural old fields
Figured modified from Keever 1950.
Recent Research on Succession
Recently, ecologists have been developing mathematical models to better characterize and predict successional changes. For example, The Resource Ratio Hypothesis, proposed by David Tilman (1985), models successional shifts in plant communities based on the assumption that succession is driven by a tradeoff in competition for nutrients in early succession, and for light in late succession. Other researchers, such as Henry Horn (1974) have used transition matrix models and Markovian models to measure rates of succession and predict the outcomes of succession.
Other current research focuses on soil organisms and foodwebs in successional systems. Until recently, there has been surprisingly little characterization of diversity patterns in non-plant organisms across successional gradients, and even less research has been devoted to the roles that these organisms may actually play in regulating succession. For example, mycorrhizal fungi, which can form mutualisms with plants, are known to play an important role in plant survival and growth, but very little is known about whether mycorrhizae can control or alter successional trajectories. In systems that initially lack mycorrhizae, such as sand dunes, only non-mycorrhizal pioneer plants are able to colonize. As mycorrhizal diversity increases in the soil, later successional plants are able to colonize (Koske & Gemma 1997). If initial soil mycorrhizal conditions vary, successional outcomes may also vary. Additional work is also being conducted on the role of soil bacteria, insects, birds, and mammals in successional processes.
The basics of successional theory are also being applied to the relatively new fields of restoration ecology and invasion biology. Restoration ecology can be viewed as an attempt to speed successional processes to reach a desired climax community. For example, prairie restoration tries to recreate a prairie climax community within 10 years, when this process naturally takes several hundred years. Restoration managers can manipulate mechanisms of succession to rapidly achieve climax conditions by greatly increasing seed availability, reducing competition by early-successional species, and amending soil to better match late-succession conditions. While not always successful, restoration efforts are often seen as an acid-test for our understanding of succession (Young 2005).
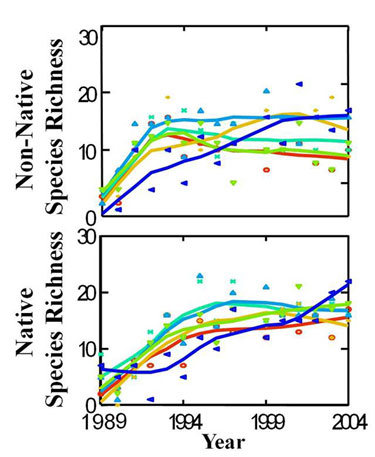
Similarly, invasive species can be viewed as a natural component of succession. The process of invasion, whereby any new species, native or non-native, is introduced and becomes established in a community, is an essential part of succession. Our understanding of successional processes can inform our understanding of the process of invasion by non-native species, at least during the initial stages of an invasion (Sakai et al. 2001). For example, in old field plant communities in the Midwestern United States, native and non-native plant diversity both increased over a period of 15 years since field abandonment (Figure 6). This indicates that invasive plants may enter new communities through processes very similar to native species. The negative impacts of non-native species are made apparent when they alter the normal successional trajectory of a system.
Figure 6: Plant diversity increases throughout succession
Native and non-native plant species richness increase over time in old fields at the Kellogg Biological Station LTER (http://lter.kbs.msu.edu/). Fields were last plowed in 1988. Different line colors indicate different field replicates (Gross & Emery 2007).
The basics of successional theory are also being applied to the relatively new fields of restoration ecology and invasion biology. Restoration ecology can be viewed as an attempt to speed successional processes to reach a desired climax community. For example, prairie restoration tries to recreate a prairie climax community within 10 years, when this process naturally takes several hundred years. Restoration managers can manipulate mechanisms of succession to rapidly achieve climax conditions by greatly increasing seed availability, reducing competition by early-successional species, and amending soil to better match late-succession conditions. While not always successful, restoration efforts are often seen as an acid-test for our understanding of succession (Young 2005).
Similarly, invasive species can be viewed as a natural component of succession. The process of invasion, whereby any new species, native or non-native, is introduced and becomes established in a community, is an essential part of succession. Our understanding of successional processes can inform our understanding of the process of invasion by non-native species, at least during the initial stages of an invasion (Sakai et al. 2001). For example, in old field plant communities in the Midwestern United States, native and non-native plant diversity both increased over a period of 15 years since field abandonment (Figure 6). This indicates that invasive plants may enter new communities through processes very similar to native species. The negative impacts of non-native species are made apparent when they alter the normal successional trajectory of a system.
Summary
While the process of succession has been studied by ecologists since the turn of the 20th century, it is still very much a dynamic field of study today. Multiple, complex mechanisms can all interact to result in predictable patterns of change in communities over time. Recently, ecologists have used principles of succession to inform the applied ecological fields of Restoration Ecology and Invasion Biology.
References and Recommended Reading
Bazzaz, F. A. Physiological ecology of plant succession. Annual Review of Ecology and Systematics 10, 351-371 (1979).
Chapin, F. S., Walker, L. R. et al. Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska. Ecological Monographs 64, 149-175 (1994).
Clements, F. E. Nature and structure of the climax. Journal of Ecology 24, 252-84 (1936).
Connell, J. H. & Slayter, R. O. Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist 111, 1119-1144 (1977).
Cooper, W. S. The recent ecological history of Glacier Bay, Alaska: the present vegetation cycle. Ecology 4, 223-246 (1923).
Cowles, H. C. The ecological relations of the vegetation on the sand dunes of Lake Michigan. Botanical Gazette 27, 95-117, 167-202, 281-308, 361-391 (1899).
Gleason, H. A. The individualistic concept of the plant association. Bulletin of the Torrey Botanical Club 53, 7-26 (1926).
Grime, J. P. Plant strategies and vegetation processes. New York, NY: John Wiley and Sons, 1979.
Horn, H. S. The ecology of secondary succession. Annual Review of Ecology and Systematics 5, 25-37 (1974).
Hubbell, S. P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, NJ: Princeton University Press, 2001.
Huston, M. & Smith, T. Plant succession- life history and competition. American Naturalist 130,168-198 (1987).
Keever, C. Causes of succession on old fields of the Piedmont, North Carolina. Ecological Monographs 20,229-250 (1950).
Koske, R. E., & Gemma, J. N. Mycorrhizae and succession in plantings of beachgrass in sand dunes. American Journal of Botany 84, 118-130 (1997).
Odum, E. P. The Strategy of Ecosystem Development. Science 164, 262-270 (1969).
Reiners, W. A., Worley, I. A. et al. Plant diversity in a chronosequence at Glacier Bay, Alaska. Ecology 52, 55-69 (1971).
Sakai, A. K., Allendorf, F. W. et al. The population biology of invasive species. Annual Review 32, 305-332 (2001).
Tilman, G. D. The resource-ratio hypothesis of succession. American Naturalist 125, 827 (1985).
Young, T. P., Petersen, D. A. et al. The ecology of restoration: historical links, emerging issues and unexplored realms. Ecology Letters 8, 662-673 (2005).










