


















« Prev Next »
The practice of biological classification appeared early in the evolution of life: Is this safe to eat/not safe to eat? Is that a predator/not a predator? Are these potential mates/not potential mates? It is in our nature, then, to classify our surroundings. Within the context of biology this practice takes on special importance, as scientists from many biological disciplines work toward a single classification system that incorporates all organisms that have ever lived. Both the practice and the product of this grand biological classification are called taxonomy.
The most widely used taxonomic system was formalized by Carolus Linnaeus and comprises a simple nested hierarchy wherein similar organisms group together at one taxonomic rank, and those groups cluster into successively broader groups at higher ranks. Figure 1 shows a taxonomy of human ancestors and their extant African ape relatives. This nested hierarchy allows different degrees of similarity to be represented at different ranks. Note, however, that Linnaean taxonomy predates modern evolutionary theory; whereas Linnaeus organized living organisms according to different levels of similarity, it was Darwin and others who explained these differing degrees of similarity through common ancestry. The more closely related two groups are, the more similarities they are likely to share. The series of evolutionary relationships among a group of organisms is termed phylogeny.

Figure 1: Taxonomy for human ancestors.
Linnaean ranks are listed across the top, with vertical lines to indicate the indentation of corresponding taxon names; ranks and taxa are also correlated by color. Ranks above the genus level include the proper taxon name (always starting with a capital letter) as well as the common name in parentheses. Hence, to refer to the group of African apes and humans, one could say either the Homininae or the hominines. The genus and species names (commonly called the "scientific name") are always written in italics, and the genus starts with a capital letter whereas the species is always lowercase. Note that the species name is never written alone, and thus each species is preceded by an abbreviation for its genus. Although nearly all workers recognize the close relationship between Homo and Pan, to the exclusion of Gorilla, there is not yet a broadly used taxonomic rank to demarcate this association.
Courtesy of Kieran P. McNulty
The Relationship Between Taxonomy and Phylogeny
It is commonly understood that taxonomy should reflect phylogeny - organisms should be grouped according to their evolutionary history, their relatedness. This makes intuitive sense because common ancestry is the only factor that unites four billion years of life on Earth. This connection between taxonomy and phylogeny means that new discoveries or other data that change our understanding of evolutionary history often result in the shuffling of taxa and of taxonomic names.
An excellent example of this shuffling is the change in usage of the term "hominid." Traditionally, only human ancestors were placed in the family Hominidae (and thereby referred to as hominids). This reflected a view that humans are substantially different from the great apes, which were placed in the Family Pongidae (pongids) (Figure 2a). However, overwhelming genetic evidence has since demonstrated that humans, chimpanzees, and gorillas are much more closely related to each other than to the orangutan (e.g., Sarich, 1971; Caccone & Powell, 1989; Ruvolo, 1994). Thus, there is no genetic support for grouping the great apes together in a distinct group from humans. For this reason, many researchers now place all species of great ape and human within a single family, Hominidae - making them all proper "hominids" (Figure 2b).
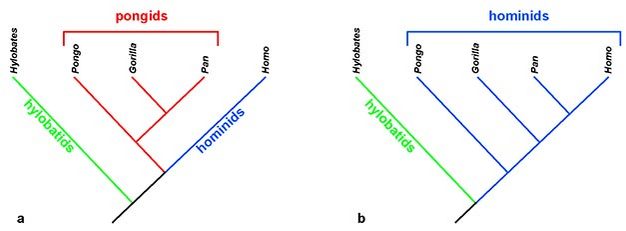
Figure 2: Changing phylogeny of the apes and implications for taxonomy.
(a) A traditional phylogeny of modern apes, where orangutans (Pongo), gorillas (Gorilla), and chimpanzees (Pan) were thought to be very closely related. In this scheme the lesser apes, gibbons and siamangs (Hylobates), are put into their own family of hylobatids; great apes are grouped together in the Pongidae (pongids), and only the human lineage was included in the Family Hominidae, the hominids. (b) This is a modern understanding of ape phylogeny, where the great apes no longer represent a distinct group from humans. Here, there is no support for a separate Family Pongidae and therefore great apes and their ancestors are grouped within the Family Hominidae. For this reason, the term "hominid" is now typically applied to all great ape and human species not merely the lineage of humans.
Courtesy of Kieran P. McNulty
Such taxonomic changes have downstream effects as well: African apes and humans are now distinguished from orangutans at the subfamily rank Homininae (hominines), and the human lineage is separated at an even lower rank of Tribe Hominini. Thus, the common term for the lineage of fossil and modern humans is "hominin." Furthermore, the traditional term "australopithecine," which grouped Australopithecus and Paranthropus in the subfamily Australopithecinae, becomes invalid under this revised taxonomy; in the context of Linnaean taxonomy, one cannot nestle a higher-ranked subfamily (Australopithecine) within a lower-ranked tribe (Hominini) (see Figure 1).
On one hand, this change in terminology over the years illustrates the proper advancement of science, whereby new evidence about evolutionary history helps to refine our taxonomic models. However, the necessary linguistic shift has been slow to gain acceptance among some researchers. As well, the popularity of terms like “hominid” and “australopithecine,” which seeped into public awareness in the 1960s-70s, means these terms will likely persist in their traditional meanings for communicating with the public.
Hominin Taxonomy and Phylogeny
One model of hominin evolution is illustrated by the phylogeny in Figure 3, presented here as a vehicle for discussion rather than a robust hypothesis. An obvious point of contention is the status of the earliest hominins, known from only small isolated samples; relationships both among these species and to the later hominins are not yet resolved. And, while other branches of the human evolutionary tree are vigorously debated in the literature, the usual signposts of phylogenetic uncertainty (e.g., dashed lines, alternate branches, question marks) have been omitted from Figure 3 for simplicity. Instead, a few of these issues are highlighted below, specifically in the context of taxonomic and phylogenic obstacles: ancestry, variation, and biological complexity.
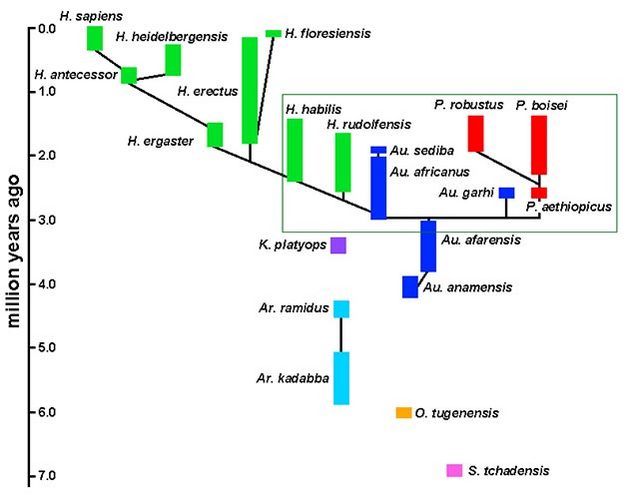
Figure 3: A phylogeny of hominin species.
Approximate geological age ranges are included for each. Black lines indicate a phylogenetic connection, though these relationships are proposed primarily for discussion and are not well-tested hypotheses. Several species are left unconnected due to the many possible ways in which they might related to the other species. The box highlights a section of the phylogeny that is used in Figure 3.
Courtesy of Kieran P. McNulty
Australopithecus and the problem of ancestry
Five species of Australopithecus are recognized here, though new discoveries are likely to add to this number. There is good evidence that A. anamensis and A. afarensis represent evolution within a broad lineage (Kimbel et al., 2006), but other relationships in the genus are not well understood. It may be that the genus is an amalgamation of stem species that are not closely related (see below), though they seem to share a common adaptive grade. Most researchers would agree that both Paranthropus and Homo evolved from some Australopithecus-like ancestor(s), though which is still contentious. This idea of ancestry, however, presents difficulties for our efforts to match taxonomy to phylogeny. For example, if the phylogeny in Figure 3 were correct, then A. africanus and its descendants would be more closely related to the genus Homo than other species of Australopithecus; likewise, A. garhi would be more closely related to Paranthropus. How, then, can our taxonomy be made to reflect these evolutionary relationships?
The most common solution is through splitting. For example, Figure 4a illustrates the cladogenetic relationships among the boxed species in Figure 3, with "Australopithecus" species here dispersed among three genera. Because the genus name Australopithecus is specifically associated with Au. africanus, this species and its descendant Au. sediba retain that genus name. "A." garhi is transferred to Paranthropus to reflect that relationship, while "A." afarensis requires a new genus designation, Praeanthropus (see, e.g., Strait et al., 2007). This horizontal splitting across the species-level of our taxonomy also implies a vertical splitting at higher ranks, where each node (junction of branches) on our cladogram requires a distinct rank to denote a separate evolutionary relationship. Thus, to exactly replicate the relationships shown in the cladogram would require adding six additional ranks (and their attendant taxon names) between the tribe and species levels representing the historical splitting events in this lineage. In this way, a taxonomic hierarchy can be made to precisely reflect the cladogenetic relationships of evolutionary history.
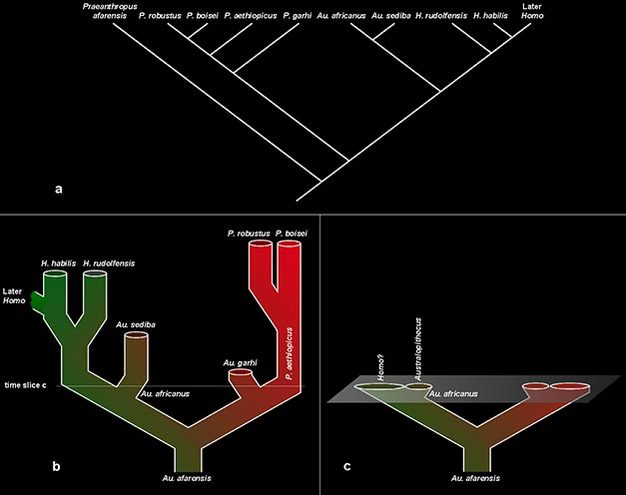
Figure 4: Systematic relationships among the hominin species
(a) A cladogram depicting possible phylogenetic relationships among these species, and the new genus designations (cf. Figure 1) that result; (b) evolutionary diagram of the same species, but depicted as a series of evolving populations rather than cladogenetic splits; the horizontal line indicates the slice of time represented in (c); (c) a time slice from the phylogeny in (b) showing the relationships between populations just after they begin to diverge.
Courtesy of Kieran P. McNulty
This approach leads to a curious theoretical dilemma, however. By arranging our taxonomic names to match phylogenetic relationships, we end up organizing species according to events that happened after their existence rather than their contemporaneous biology. Consider, for example, the phylogeny in Figure 4b, depicting these populations evolving through time. Considering a time slice from this phylogeny (Figure 4c), the two populations of Au. africanus would be - by any contemporaneous measure - the same species. But, because one population later evolved into species of Homo, researchers at a modern vantage are inclined to transfer that population into this genus. Such action might better reflect the subsequent phylogenetic relationships among descendants, but does not fully account for the biological context of the organisms in question.
Early Homo and the problem of variation
The divide between australopiths and the genus Homo was once fairly clear, the latter group distinguished by a markedly larger brain. Leakey et al.'s (1964) addition to Homo of the smaller-brained H. habilis, however, blurred this boundary and sparked debate over the scope of variation that could be accommodated within the human genus. With the discovery of many more specimens of early Homo, however, these arguments gave way to other taxonomic considerations: how many species were represented in the diverse sample of small-brained Homo? could they all be accommodated within the single species H. habilis?
Among fossils, variation within a species is largely a taxonomic issue rather than a phylogenetic one simply because fossil preservation is rarely good enough to develop robust evolutionary hypotheses for a large number of individual specimens. Unfortunately, there are no clear guidelines as to the amount of variation appropriate to any taxonomic group, and no definitive way to test whether species allocations are appropriate. Researchers commonly use variation within modern species as benchmarks for acceptable ranges and patterns of variation (e.g., McNulty, 2003, 2005; Baab et al., 2010), but even this can be problematic: variation in extinct species need not be similar to variation in modern ones (Kelley, 1993). This problem may be especially acute in hominin studies, where variation in living comparator species, such as gorillas and chimpanzees, may have been radically impacted by declining populations.
What, then, is the nature of variation in early Homo? A number of comparisons to modern apes and humans have found that the sample traditionally grouped in H. habilis exceeded the variation expected within a single modern species (e.g., Stringer 1986; Bilsborough and Wood 1988; Lieberman et al. 1988; Chamberlain 1989; Kramer et al. 1995). This led Alexeev (1986) to propose a new species, H. rudolfensis, to accommodate the larger specimens of this sample, retaining the smaller ones in H. habilis. With one large and one small species of early Homo, the logical counter argument was that the sample represents a single, sexually dimorphic species (e.g., Howell 1978); Wood et al. (1991), however, provided tentative evidence that patterns of variation within the broad sample of early Homo are not consistent with known patterns of dimorphism. For this reason, paleoanthropologists have largely accepted the presence of two species of early Homo, and there may be evidence for additional taxonomic diversity (Smith and Grine, 2008).
As a final twist in the taxonomic narrative, Wood and Collard (1999) provided a coherent justification for removing H. habilis and H. rudolfensis from Homo entirely, and placing one or both into Australopithecus (see also Leakey et al., 2001). Thus far, however, their proposal has not gained broad acceptance.
Neanderthals and the problem of biological complexity
One of the oldest debates in paleoanthropology revolves around the taxonomic status of the neanderthals. While most researchers agree that the distinctive neanderthal morphology indicates a degree of separation from other contemporary hominins, there is disagreement over whether neanderthals belong within H. sapiens or in their own species, H. neanderthalensis. There is ample morphological justification for recognizing a distinct species (e.g., Harvati et al., 2004), and early analyses of ancient mitochondrial DNA from neanderthals supported this conclusion (e.g., Krings et al., 1997). More recently, however, sequencing of the nuclear genome has shown that modern human populations outside of Africa retain up to 4% of distinctive neanderthal genes (Green et al., 2010). If one defines a species as populations of interbreeding organisms (a criterion known as the biological species concept), then the presence of neanderthal genes among modern humans argues strongly for including them within H. sapiens, because it implies that contemporaneous neanderthals and anatomically modern humans were interbreeding.
The case of the neanderthals demonstrates a final hurdle in the standard practice of taxonomy and phylogeny: biology does not always adhere to our simplistic models. Our preference for arbitrating discrete taxonomic groups and instantaneous phylogenetic splits ignores the continuity and complexity of biological evolution. Indeed, one of the most compelling demonstrations of evolution and common ancestry is precisely the lack of strict demarcations between closely related species: dogs and wolves interbreed; different monkey species hybridize; even the mule (or hinny) can produce offspring, albeit rarely. Such blurred boundaries are exactly what would be expected from populations that continuously evolve and diverge over time. This biological reality, however, plays havoc with notions that evolutionary lineages cleave instantaneously, or that we can perfectly circumscribe groups of organisms into discrete taxonomic groups.
Did neanderthals split into a separate species or were they part of our direct lineage? The answer is probably "both." Reproductive isolation, and thereby speciation, is a process through which populations experience degrees of infertility - behavioral and structural impediments, reduced hybrid viability, gametic incompatability, etc. - over many generations. In this sense, lineages may not cleave so much as reticulate, with the network of genetic exchange between populations dissipating over time. Even in neanderthals, who were likely geographically isolated from the modern human lineage, the capacity to interbreed was persistent over hundreds of thousands of years (Green et al., 2010). Nevertheless, with neanderthal DNA providing perhaps the only opportunity to directly test whether an extinct pre-modern hominin interbred with modern H. sapiens, I am inclined here to group neanderthals within our species.
The Importance of Taxonomy in Paleoanthropology
We study human ancestry not simply to have labels for museum exhibits, but rather to illuminate the complex story of morphological, physiological, behavioral, cognitive, and cultural changes that gave rise to modern humans. Thus, taxonomy in itself is not of direct interest. Coupled with its primary use as a substitute for phylogeny, taxonomy may seem easy to disregard. Indeed, if one worked with only modern species - who have complete anatomies, DNA sequences, behaviors - it might be possible to rely entirely on phylogenetic relationships to organize specimens into meaningful groups.
Not so in the study of fossils, however, where finding any remnant of ancient species is rare, and finding pieces that are relatively intact is extraordinary. Lacking sufficient anatomy for robust phylogenetic analyses of each bone fragment in a study, it is taxonomic hypotheses that form the biological basis for organizing fossil samples. Within paleoanthropology, then, taxonomy still plays three crucial roles:
1. Alpha-taxonomy. The sorting of individual specimens into species is perhaps the most important role of taxonomy in modern paleoanthropology. In a sample of bone fragments that includes different pieces of anatomy as well as individuals of different size, age, sex, geological antiquity, and perhaps species, it is alpha-taxonomy that provides the justification for uniting some specimens into groups while separating others - usually through reference to robust extant models of within-species variation. Importantly, because nearly all paleoanthropological research is based on taxonomic hypotheses, either explicitly or implicitly, changes in the underlying taxonomy have the capacity to greatly impact our interpretation of human evolution.
2. Mediating biologically meaningful comparisons across space/time. Beyond the species level, taxonomic analyses provide a mechanism for comparing broader groups of fossil organisms. Working in the fossil record means analyzing samples from different places, different times, and known at different phylogenetic resolutions. Yet, that resolution determines the biological coherence of the phylogenetic groups: a highly resolved section of a phylogeny might include closely related fossil species whereas an unresolved section could pool a much wider range of organisms. Taxonomic hypotheses allow researchers to maintain some level of uniformity in comparisons across such large groups, regardless of the phylogenetic resolution.
3. Communication. One of the greatest impacts of Linnaeus's taxonomy was in facilitating communication in the scientific community. This is still important today, and there is currently no broadly accepted vehicle to replace it. The basic terminology of taxonomy is also recognizable to the general public, and it is through such terms that research is made widely available. If the knowledge generated through research into millions of years of human evolution is to have any impact beyond the walls of the academy, then traditional taxonomic labels will likely maintain a place in paleoanthropology.
The Myth of the Ever-Changing Hominin Phylogeny
One of the common plaints leveled against paleoanthropologists is that they are constantly re-arranging the human evolutionary tree. Indeed, it does seem that every significant new fossil is purported to "completely change our understanding of human evolution." In reality, however, hominin phylogeny has been relatively stable over the last two decades, despite significant and contentious new discoveries. Comparing phylogenies published in the early 1990s to those appearing in 2012 shows a remarkable level of conformity. This suggests that - at least in broad strokes - our hypotheses about the evolution of the human lineage are fairly robust.
Consider again the sample of hominin species through time, but focusing instead on the three well-sampled genera: Australopithecus, Paranthropus, and Homo (Figure 5). Substantial fossil samples from each have been accumulating for nearly a century - longer, in the case of fossil Homo. The history of studies on these specimens generates a great deal of stabilizing inertia to the basic relationships shown here. While specific connections between these groups, as well as the phylogenetic relationships within them, are still far from resolved, the overall arrangement of Australopithecus, Paranthropus, and Homo seems relatively stable; indeed, any changes to this arrangement are likely to be, well, taxonomic.
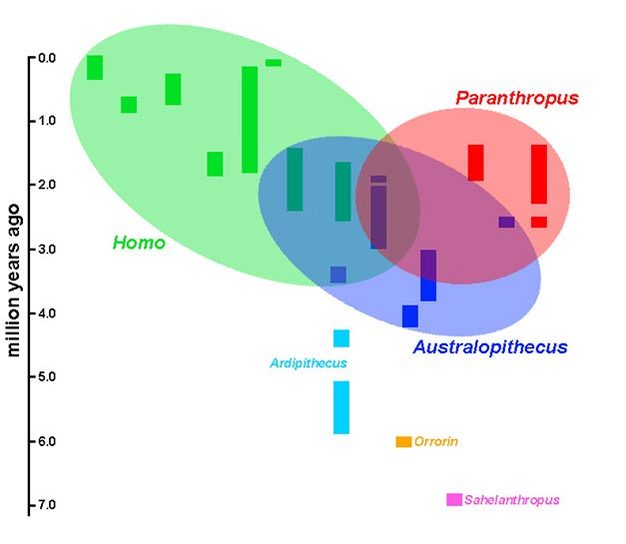
Figure 5: Relationships among hominin genera and regions of ongoing debate
A reiteration of the hominin species distributions, but emphasizing more broadly the basic relationships among Australopithecus, Paranthropus, and Homo. Each ellipse encircles the species that belong in a genus, but also extends to overlap with other species that have been implicated in debates over the genus. Thus, the regions where two or three ellipses overlap are likely to represent important areas for continuing research.
Courtesy of Kieran P. McNulty
That being said, which aspects of paleoanthropological research are most likely to have a major impact on our knowledge of human evolutionary relationships? The ready answer is from new discoveries. Findings such as the fossil "hobbits" in Flores (H. floresiensis) and the enigmatic Homo species from Dmanisi have dramatic implications for human evolution, and the significance of these has yet to fully impact hominin phylogeny. The status of the earliest hominins is also in a state of flux, and new discoveries at the base of the hominin phylogeny will surely be felt up in the higher branches. Perhaps the greatest potential for revising our interpretation of human evolution, however, comes from some of the better known taxa. Referring again to Figure 5, the ellipses have been drawn so as to encompass ongoing taxonomic and phylogenetic debates relevant to each genus. As an heuristic device, then, the regions where these ellipses overlap are likely to produce the greatest impact on our understanding of human evolution. Resolving these relationships - determining which species ultimately belong to which genera, and how those genera are related - has the potential to transform our understanding of human origins and diversification.
Glossary
Australopiths: a general term typically used in reference to species of Australopithecus and Paranthropus. This is not a formal taxonomic name and therefore does not need to adhere to the strict rules of biological nomenclature.
Biological species concept: This is the most commonly used definition of a "species," whereby populations of organisms are determined to belong to the same species if their members interbreed and produce viable offspring.
Cladogenetic: adjective referring to evolution through splitting events. If a population diverges into two branches which thereafter evolve independently of one another, this is called cladogenesis (or, cladogenetic evolution). If a population evolves through time without splitting into multiple branches it is called anagenesis (or, anagenetic evolution).
Cladogram: a branching diagram that represents only phylogenetic relationships among organisms (i.e., it does not include geological age or other contextual information). Formally, a cladogram is produced through cladistic analysis of a group of organisms, which determines evolutionary relationships using only derived (evolved) characteristics that are shared among at least some members of a group.
Common ancestry: the shared evolutionary history of all living organisms back to a single origin.
Gametic incompatibility: a barrier to sexual reproduction where the sex cells (gametes) of a male (sperm) and female (egg) are too different to allow fertilization. Once two populations have diverged and begin evolving independently, eventually their members will become incapable of interbreeding due to gametic incompatibility. One example of this might be if evolution results in different numbers of chromosomes in the two populations so that their members cannot interbreed (note that different chromosome numbers does not always lead to gametic incompatibility).
Mitochondrial DNA: genetic information that is associated with a cell's mitochondria rather than its chromosomes. The mitochondria resides outside of the cell nucleus (within the cytoplasm) and are inherited only from the maternal line in sexually reproducing organisms.
Node: Evolutionary relationships are often represented by tree-like patterns of branching lineages. A node is where two or more branches come together at a point, and hence it represents the last common ancestor shared by those groups before they diverged.
Nuclear genome: The nuclear genome is made up of the DNA from an organisms chromosomes, which reside in the cell nucleus. In sexually reproducing creatures, the nuclear genome contains genes from both parents.
Phylogeny: the evolutionary relationships among, or evolutionary history of, a group of organisms.
Sexually dimorphic: Adjectival phrase referring to differences in size and shape between males and females of a species.
Taxon: Any group from any rank of the Linnaean classification can be generally referred to as a taxon. Thus, the Family Hominidae is a taxon, as is the species H. sapiens and the Order Primates. The plural is taxa.
Taxonomic rank: This refers to different levels of the Linnaean hierarchical classification. For example, species is one taxonomic rank; kingdom, phylum, and class are other ranks.
Taxonomy: the classification of living organisms, typically using the hierarchical system invented by Carolus Linnaeus.
References and Recommended Reading
Alexeev, V. P. The Origin of the Human Race. Moscow: Progress Publishers, 1986.
Baab, K. L., McNulty, K. P., et al. A comprehensive comparison of the Homo floresiensis cranium to pathological humans and extinct hominins. PLOS One 8, e69119.
Bilsborough, A., & Wood, B. A. Cranial morphometry of early hominids I. Facial region. American Journal of Physical Anthropology 76, 61-86 (1988).
Caccone, A., & Powell, J. R. DNA divergence among hominoids. Evolution 43, 925-942 (1989).
Chamberlain, A.T. Variations within Homo habilis. In Hominidae: Proceedings of the 2nd International Congress of Human Paleontology. ed. Giacobini, G. (Milan: Jaca Books, 1989). 175-181.
Green, R. E., Krause, J., et al. A draft sequence of the neandertal genome. Science 328, 710-722 (2010).
Harvati, K., Frost, S. R., et al. Neanderthal taxonomy reconsidered: Implications of 3D primate models of intra- and interspecific differences. Proceedings of the National Academy of Sciences, USA 101, 1147-1152 (2004).
Howell, F. C. Hominidae. In Evolution of African Mammals. eds. Maglio, V. J., & Cooke, H.B.S. (Cambridge: Harvard University Press, 1978). 154-258.
Kelley, J. Taxonomic implications of sexual dimorphism in Lufengpithecus. In Species, Species Concepts and Primate Evolution. eds. Kimbel, W. H. & Martin, L. B. (New York: Plenum Press, 1993). 429-458.
Kimbel, W. H. , Lockwood, C. A., et al. Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record. Journal of Human Evolution 51, 134-152 (2006).
Kramer, A., Donnelly, S. M., et al. Craniometric variation in large-bodied hominoids: testing the single-species hypothesis for Homo habilis. Journal of Human Evolution 29, 443-462 (1995).
Krings, M., Stone, A., et al. Neandertal DNA sequences and the origin of modern humans. Cell 90, 19-30 (1997).
Leakey, L. S. B., Tobias, P. V., et al. New species of the genus Homo from Olduvai Gorge. Nature 202, 7-9 (1964).
Leakey, M. G., Spoor, F., et al. New hominin genus from Eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433-440 (2001).
Lieberman, D. E., Pilbeam, D. R., et al. A probabilistic approach to the problem of sexual dimorphism in Homo habilis: a comparison of KNM-ER 1470 and KNM-ER 1813. Journal of Human Evolution B, 503-511 (1988).
McNulty, K. P. Geometric morphometric analyses of extant and fossil hominoid craniofacial morphology. Unpubl. Ph.D. thesis. City University of New York, New York (2003).
McNulty, K. P. A geometric morphometric assessment of the hominoid supraorbital region: Affinities of the Eurasian Miocene hominoids Dryopithecus, Graecopithecus, and Sivapithecus. In Modern Morphometrics in Physical Anthropology. ed. Slice, D. (New York: Kluwer Academic Publishers, 2005). 349-373.
Ruvolo, M. Molecular evolutionary processes and conflicting gene trees: the hominoid case. American Journal of Physical Anthropology 94, 89-113 (1994).
Sarich, V. M. A molecular approach to the question of human origins. In Background for Man. ed. Dolhinow, P. (Boston: Little, Brown, 1971). 60-81.
Smith, H. F., & Grine, F. E. Cladistic analysis of early Homo crania from Swartkrans and Sterkfontein, South Africa. Journal of Human Evolution 54, 684-704 (2008).
Strait, D., Grine, F. E. et al. Analyzing hominid phylogeny. In Handbook of Paleoanthropology, vol. 3. eds. Henke, W., & Tattersall, I. ( Berlin: Springer, 2007). 1781-1806.
Stringer, C. B. The credibility of Homo habilis. In Major Topics in Primate and Human Evolution . eds. Wood, B. A., Martin, L. B., et al. (Cambridge: Cambridge University Press, 1986). 266-294.
Wood, B. A., & Collard, M. The changing face of the genus Homo. Evolutionary Anthropology, 8, 195-207 (1999).
Wood, B. A., Yu, L., et al. Intraspecific variation and sexual dimorphism in cranial and dental variables among higher primates and their bearing on the hominid fossil record. Journal of Anatomy 174, 185-205 (1991).










