


















« Prev Next »


Figure 1: Alfred Russel Wallace, Father of Biogeography
This article describes some of the major geographic patterns in species richness, and the processes and theories that are thought to account for these patterns. Much of this knowledge has emerged from the tremendous body of work from one scientist, Alfred Russel Wallace (Figure 1), widely regarded as the “Father of Biogeography.” Aside from co-originating the process of Natural Selection with Charles Darwin, Wallace spent extended periods studying the distribution and diversity of plants and animals in Amazonia and Southeast Asia in the mid 1800s. Many of the patterns and processes featured in this article were initially described by Wallace, and careful study of his work indicates that his ideas presaged many of the discoveries made by his numerous successors.
Many of the spatial patterns in biodiversity are overt, others are subtle and yet additional patterns remain undetected. While the existence of these patterns may be obvious — and changes in the environment that are paired with these patterns may also be obvious — the mechanisms that cause the differences in biodiversity along environmental gradients are under still the subject of scientific debate. Because large-scale patterns are the emergent result of complex interactions at many spatial and temporal scales, no single answer is likely to ever emerge, but with continued research our understanding of the processes shaping these patterns increases.
Historical Processes Affecting Biogeography
Figure 2: Darwin's finches of the Galapagos Islands are a classic example of adaptive radiation.
The study of historical factors shaping species richness and distribution is often broken into two major categories: vicariance and disperal. Species can occur in a location because their ancestors remained there passively as the environment moved around them (vicariance). Alternatively, a species or its ancestors may have arrived at a location via movement from another location (dispersal). Vicariance describes the disruption of the biogeographic range of a group of organisms by changes in the environment. Vicariant events can happen when landmasses move apart through tectonic action, or when mountains emerge to divide the geographic ranges of species. Vicarance typically leads to the emergence of new species through allopatric speciation, in which one ancestral species will result in the production of two new species that evolve apart from one another in geographic isolation, frequently by genetic drift rather than natural selection. Dispersal biogeography focuses on the movement of species from one location to another location. Jump dispersal events, when individuals of a species travel a relatively long distance to a new environment in which they did not previously occur, can result in the adaptive radiation of one ancestral species giving rise to a broad diversity of new species. Classic examples of adaptive radiation include fruit flies in the Hawaiian Islands, finches on the Galapagos Islands (Figure 2), and anolis lizards throughout Central and South America and the Caribbean. Both the Galapagos and Hawaiian islands emerged from the sea floor in the ocean and all species occurring on such oceanic islands needed to arrive via dispersal, whereas continental islands which broke off from a mainland may contain species as a result of vicariance.
Major Spatial Patterns in Biodiversity
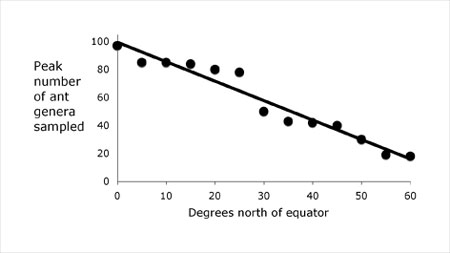
One major geographic pattern in biodiversity is the latitudinal gradient in species richness. As one travels further away from the equator, for most taxa, the number of species declines. For example, Figure 3 demonstrates the latitudinal richness gradient for richness in ants of the Northern Hemisphere. This general pattern holds true for most taxa and ecosystem types in both marine and terrestrial environments. There is broad agreement that this pattern is caused by differences in the abiotic, climatic environment, but the specific mechanism or mechanisms causing this pattern are a continued topic of discussion and investigation. One set of theories, broadly grouped together as “species-energy theory” is based on the fact that the amount of radiant energy from the sun captured by ecosystems is negatively associated with latitude. As energy is distributed throughout ecosystems through trophic processes, it is thought that species richness will track the energy following one or more mechanisms. Models of species-energy theory incorporate variables such as temperature, net primary productivity, speciation, and extinction. Other ideas that have been proposed to account for the latitudinal gradient are related to the physiological responses of animals to climatic conditions and the effects of the abiotic environment on historical processes. Most of these theories are not mutually exclusive.
Another recurring pattern in biogeographic theory is the elevational gradient in species richness. As one travels to higher elevations, the number of species declines, or, in many cases, peaks at mid-elevations. Aside from the environmental mechanisms driving this diversity gradient, there is a phenomenon that is based on the geography of species range distributions called the mid-domain effect. The mid-domain effect predicts a peak of diversity at the midpoint along any domain simply by the fact that the ranges of more species overlap in the middle of a domain (like a mountain or an island) than on the edges, and this effect works together with environmental determinants to affect the net distribution of species along many elevational gradients.
A third recurring pattern in the distribution of species is the area effect on species richness. The larger an place is, the more species it can support. This applies to actual islands in bodies of water, as well as habitat islands such as those surrounded by human development. The species-area relationship is presented in more detail in an article about the theory of island biogeography.
Figure 3: The latitudinal gradient in richness in ants
The maximum number of genera sampled decreases as one moves north from the equator.
Another recurring pattern in biogeographic theory is the elevational gradient in species richness. As one travels to higher elevations, the number of species declines, or, in many cases, peaks at mid-elevations. Aside from the environmental mechanisms driving this diversity gradient, there is a phenomenon that is based on the geography of species range distributions called the mid-domain effect. The mid-domain effect predicts a peak of diversity at the midpoint along any domain simply by the fact that the ranges of more species overlap in the middle of a domain (like a mountain or an island) than on the edges, and this effect works together with environmental determinants to affect the net distribution of species along many elevational gradients.
A third recurring pattern in the distribution of species is the area effect on species richness. The larger an place is, the more species it can support. This applies to actual islands in bodies of water, as well as habitat islands such as those surrounded by human development. The species-area relationship is presented in more detail in an article about the theory of island biogeography.
Do Neutral Models Explain Patterns in Species Richness?

Figure 4: Cane toad (Rhinella marina) is an invasive species.
We have also talked about large-scale effects on diversity — such as climate and area, but do local interactions – like competition and predation — influence the distribution of biodiversity? The fact that some species have greater effects on species richness than others may be gleaned from studying the biology of invasive species. An invasive species is one which is transported outside its original geographic range to a novel habitat, where it increases in density, and can cause detrimental effects on the indigenous species in that area, often reducing biodiversity. One example of an invasive species is the Argentine ant (Linepithema humile); more than 100 years ago this species was transported from Argentina to most other biogeographic regions, and where this species occurs the local species of other ants is markedly reduced. Another example of an invasive species with negative effects on native faunas are cane toads (Rhinella marina) introduced from the Neotropics into Australia in 1935 (Figure 4). Cane toads have reduced prey availability for indigenous predators and caused declines in native Australian amphibians. Most introduced species do not become invasive, indeed they frequently become extinct because they are less likely to be well adapted to the new environment than indigenous species, but the subset of introduced species that become invasive is problematic in terms of the economy, ecosystem services, and biodiversity. The widespread exchange of species among distant parts of the world as a result of human commerce has clearly resulted in a loss of biodiversity at many spatial scales.
Summary
The structure of the world is predictive of the species richness. Factors that predict species richness include the size and position of continentals and islands, the height and position of mountains, and temperature and access to energy from the sun. Moreover, the effects of geography on species richness are not limited to conditions we currently observe, as many of the patterns we observe are contingent on historic movements of land masses and the uplift and erosion of mountains, and the associated effects on climate and the persistence and dispersal of organisms. By studying how richness varies from location to location, we more generally understand its causes and better predict its future.
References and Recommended Reading
Colwell, R. K. & Lees, D. C. The mid-domain effect: geometric constraints on the geography of species richness. Trends in Ecology & Evolution 15, 70-76 (2000).
Hillebrand, H. On the generality of the latitudinal diversity gradient. The American Naturalist 163, 192-211 (2004).
Hubbell, S. P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, NJ: Princeton University Press, 2001.
Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecology Letters 8, 224-239 (1995).
Wallace, A. R. The Malay Archipelago. London, UK: Macmillan and Company, 1869.










