


















« Prev Next »

Nitrogen is a critical limiting element for plant growth and production. It is a major component of chlorophyll, the most important pigment needed for photosynthesis, as well as amino acids, the key building blocks of proteins. It is also found in other important biomolecules, such as ATP and nucleic acids. Even though it is one of the most abundant elements (predominately in the form of nitrogen gas (N2) in the Earth’s atmosphere), plants can only utilize reduced forms of this element. Plants acquire these forms of “combined” nitrogen by: 1) the addition of ammonia and/or nitrate fertilizer (from the Haber-Bosch process) or manure to soil, 2) the release of these compounds during organic matter decomposition, 3) the conversion of atmospheric nitrogen into the compounds by natural processes, such as lightning, and 4) biological nitrogen fixation (Vance 2001).
Biological nitrogen fixation (BNF), discovered by Beijerinck in 1901 (Beijerinck 1901), is carried out by a specialized group of prokaryotes. These organisms utilize the enzyme nitrogenase to catalyze the conversion of atmospheric nitrogen (N2) to ammonia (NH3). Plants can readily assimilate NH3 to produce the aforementioned nitrogenous biomolecules. These prokaryotes include aquatic organisms, such as cyanobacteria, free-living soil bacteria, such as Azotobacter, bacteria that form associative relationships with plants, such as Azospirillum, and most importantly, bacteria, such as Rhizobium and Bradyrhizobium, that form symbioses with legumes and other plants (Postgate 1982). These organisms are summarized in Figure 1.
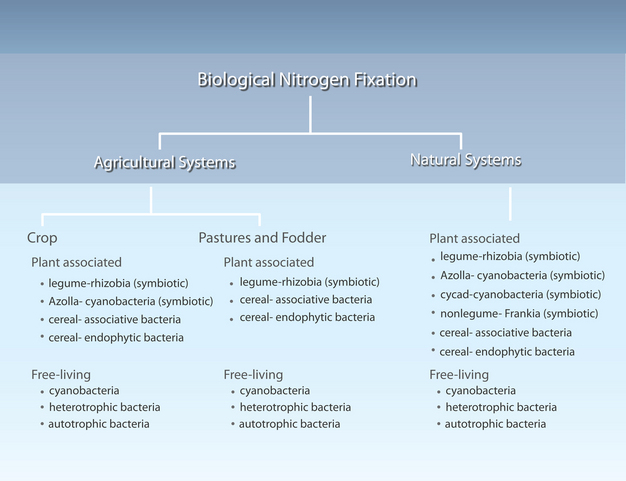
Figure 1
Nitrogen-fixing organisms found in agricultural and natural systems.
The Process
The reduction of atmospheric nitrogen is a complex process that requires a large input of energy to proceed (Postgate 1982). The nitrogen molecule is composed of two nitrogen atoms joined by a triple covalent bond, thus making the molecule highly inert and nonreactive. Nitrogenase catalyzes the breaking of this bond and the addition of three hydrogen atoms to each nitrogen atom.
Microorganisms that fix nitrogen require 16 moles of adenosine triphosphate (ATP) to reduce each mole of nitrogen (Hubbell & Kidder, 2009). These organisms obtain this energy by oxidizing organic molecules. Non-photosynthetic free-living microorganisms must obtain these molecules from other organisms, while photosynthetic microorganisms, such as cyanobacteria, use sugars produced by photosynthesis. Associative and symbiotic nitrogen-fixing microorganisms obtain these compounds from their host plants’ rhizospheres (National Research Council 1994, Hubbell & Kidder 2009).
Industries use the Haber-Bosch process to reduce nitrogen essentially in the same way. Conventional agriculture has depended upon this process to produce the commercial fertilizer needed to grow most of the world’s hybrid crops. But this approach comes with many consequences, including using fossil fuels for the energy needed to produce this fertilizer, the resulting carbon dioxide emissions and pollution from burning these fuels, and adverse affects on human health (Vitousek 1997).
Overuse of these chemical fertilizers has led to an upset in the nitrogen cycle and consequently to surface water as well as groundwater pollution. Increased loads of nitrogen fertilizer to freshwater, as well as marine ecosystems, has caused eutrophication, the process whereby these systems have a proliferation of microorganisms, especially algae. This “greening” of the water column has caused decreased levels of dissolved oxygen (DO) in bottom waters as planktonic algae die and fuel microbial respiration. These depleted DO levels result in massive mortality of aquatic organisms and create so-called dead zones, areas where little or no aquatic life can be found (Figure 2). Since the 1960’s, dead zones have increased exponentially worldwide, and have now been documented from over 400 systems, affecting more than 245,000 square kilometers of coastal regions (Diaz & Rosenberg 2008, Figure 3). This phenomenon is now deemed the key stressor on marine ecosystems.

Figure 2
True-color image of Mississippi River sediment deposition into the Gulf of Mexico.
Courtesy of NASA.
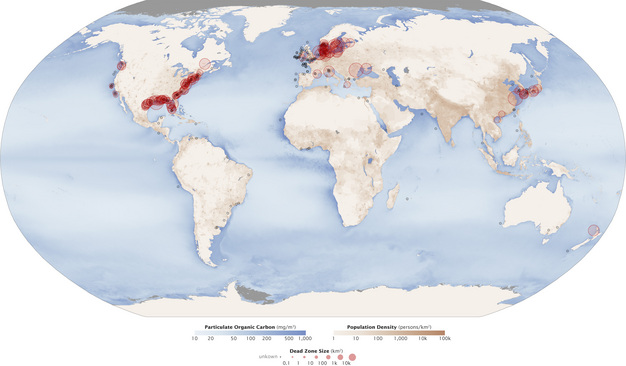
Figure 3
Aquatic dead zones across the world.
Courtesy of NASA.
Nitrogen Fixation by Free-Living Heterotrophs
Many heterotrophic bacteria live in the soil and fix significant levels of nitrogen without the direct interaction with other organisms. Examples of this type of nitrogen-fixing bacteria include species of Azotobacter, Bacillus, Clostridium, and Klebsiella. As previously noted, these organisms must find their own source of energy, typically by oxidizing organic molecules released by other organisms or from decomposition. There are some free-living organisms that have chemolithotrophic capabilities and can thereby utilize inorganic compounds as a source of energy.
Because nitrogenase can be inhibited by oxygen, free-living organisms behave as anaerobes or microaerophiles while fixing nitrogen. Because of the scarcity of suitable carbon and energy sources for these organisms, their contribution to global nitrogen fixation rates is generally considered minor. However, a recent study in Australia of an intensive wheat rotation farming system demonstrated that free-living microorganisms contributed 20 kilograms per hectare per year to the long-term nitrogen needs of this cropping system (30-50% of the total needs; Vadakattu & Paterson 2006). Maintaining wheat stubble and reduced tillage in this system provided the necessary high-carbon, low-nitrogen environment to optimize activity of the free-living organisms.
Because nitrogenase can be inhibited by oxygen, free-living organisms behave as anaerobes or microaerophiles while fixing nitrogen. Because of the scarcity of suitable carbon and energy sources for these organisms, their contribution to global nitrogen fixation rates is generally considered minor. However, a recent study in Australia of an intensive wheat rotation farming system demonstrated that free-living microorganisms contributed 20 kilograms per hectare per year to the long-term nitrogen needs of this cropping system (30-50% of the total needs; Vadakattu & Paterson 2006). Maintaining wheat stubble and reduced tillage in this system provided the necessary high-carbon, low-nitrogen environment to optimize activity of the free-living organisms.
Associative Nitrogen Fixation
Species of Azospirillum are able to form close associations with several members of the Poaceae (grasses), including agronomically important cereal crops, such as rice, wheat, corn, oats, and barley. These bacteria fix appreciable amounts of nitrogen within the rhizosphere of the host plants. Efficiencies of 52 mg N2 g-1 malate have been reported (Stephan et al. 1979). The level of nitrogen fixation is determined by several factors, including soil temperature (Azospirillum species thrive in more temperate and/or tropical environments), the ability of the host plant to provide a rhizosphere environment low in oxygen pressure, the availability of host photosynthates for the bacteria, the competitiveness of the bacteria, and the efficiency of nitrogenase (Vlassak & Reynders, 1979).
Symbiotic Nitrogen Fixation
Many microorganisms fix nitrogen symbiotically by partnering with a host plant. The plant provides sugars from photosynthesis that are utilized by the nitrogen-fixing microorganism for the energy it needs for nitrogen fixation. In exchange for these carbon sources, the microbe provides fixed nitrogen to the host plant for its growth.
One example of this type of nitrogen fixation is the water fern Azolla’s symbiosis with a cyanobacterium Anabaena azollae. Anabaena colonizes cavities formed at the base of Azolla fronds. There the cyanobacteria fix significant amounts of nitrogen in specialized cells called heterocysts. This symbiosis has been used for at least 1000 years as a biofertilizer in wetland paddies in Southeast Asia. Rice paddies are typically covered with Azolla “blooms” that fix up to 600 Kg N ha-1 yr-1 during the growing season (Postgate 1982, Fattah 2005).
Another example is the symbiosis between actinorhizal trees and shrubs, such as Alder (Alnus sp.), with the actinomycete Frankia. These plants are native to North America and tend to thrive in nitrogen-poor environments. In many areas they are the most common non-legume nitrogen fixers and are often the pioneer species in successional plant communities. Actinorhizal plants are found in many ecosystems including alpine, xeric, chapparal, forest, glacial till, riparian, coastal dune, and arctic tundra environments (Benson & Silvester, 1993).
Even though the symbiotic partners described above play an important role in the worldwide ecology of nitrogen fixation, by far the most important nitrogen-fixing symbiotic associations are the relationships between legumes and Rhizobium and Bradyrhizobium bacteria. Important legumes used in agricultural systems include alfalfa, beans, clover, cowpeas, lupines, peanut, soybean, and vetches. Of the legumes in agricultural production, soybeans are grown on 50% of the global area devoted to legumes, and represent 68% of the total global legume production (Vance 2001).
One example of this type of nitrogen fixation is the water fern Azolla’s symbiosis with a cyanobacterium Anabaena azollae. Anabaena colonizes cavities formed at the base of Azolla fronds. There the cyanobacteria fix significant amounts of nitrogen in specialized cells called heterocysts. This symbiosis has been used for at least 1000 years as a biofertilizer in wetland paddies in Southeast Asia. Rice paddies are typically covered with Azolla “blooms” that fix up to 600 Kg N ha-1 yr-1 during the growing season (Postgate 1982, Fattah 2005).
Another example is the symbiosis between actinorhizal trees and shrubs, such as Alder (Alnus sp.), with the actinomycete Frankia. These plants are native to North America and tend to thrive in nitrogen-poor environments. In many areas they are the most common non-legume nitrogen fixers and are often the pioneer species in successional plant communities. Actinorhizal plants are found in many ecosystems including alpine, xeric, chapparal, forest, glacial till, riparian, coastal dune, and arctic tundra environments (Benson & Silvester, 1993).
Even though the symbiotic partners described above play an important role in the worldwide ecology of nitrogen fixation, by far the most important nitrogen-fixing symbiotic associations are the relationships between legumes and Rhizobium and Bradyrhizobium bacteria. Important legumes used in agricultural systems include alfalfa, beans, clover, cowpeas, lupines, peanut, soybean, and vetches. Of the legumes in agricultural production, soybeans are grown on 50% of the global area devoted to legumes, and represent 68% of the total global legume production (Vance 2001).
Legume Nodule Formation
The Rhizobium or Bradyrhizobium bacteria colonize the host plant’s root system and cause the roots to form nodules to house the bacteria (Figure 4). The bacteria then begin to fix the nitrogen required by the plant. Access to the fixed nitrogen allows the plant to produce leaves fortified with nitrogen that can be recycled throughout the plant. This allows the plant to increase photosynthetic capacity, which in turn yields nitrogen-rich seed. The consequences of legumes not being nodulated can be quite dramatic, especially when the plants are grown in nitrogen-poor soil. The resulting plants are typically chlorotic, low in nitrogen content, and yield very little seed (Figure 5 and 6).



The nodulation process illustrates an orchestrated interaction between the bacteria and host plant (Napoli & Hubbell 1975, Kamst et al. van Rhyn & Vanderleyden 1995, Cheng & Walker 1998). The process begins when the rhizobia are attracted to flavonoids released by the host legume’s roots. For legumes like alfalfa, clover, and soybeans (others like lupines and peanuts form nodules in other ways) the bacteria then begin to attach themselves to extensions of root epidermal cells called root hairs. The attachment process is actually a two-step process where the bacteria first attach using a Ca2+ - binding protein called rhicadhesin. After the bacteria accumulate and anchor themselves to the root hair surface, a firmer attachment that involves lectins and/or cellulose firbrils and fimbriae produced by the host plant and bacteria, respectively.
The host legume then senses chemicals produced by the rhizobia called Nod factors that cause the colonized root hairs to curl and form what is called a shepherd’s crook. Then rhizobia penetrate the root hairs and typically form a tubular structure called an infection thread. Once the bacteria reach the root itself, they stimulate cortical cell divisions that lead to the formation of a nodule. As the nodule begins to form, the bacteria become surrounded by a plant-derived membrane and are released inside plant cells forming the nodule. The bacteria subsequently lose their cell walls and undergo a profound change in cell morphology to form large, irregularly shaped branching cells called bacteroids. They then are entirely dependent on the host plant for their energy needs. In return, the bacteria fix nitrogen for the plant.
The interaction between the bacteria and host legume is so intricate that a particular Rhizobium or Bradyrhizobium will only nodulate a select number of plant genera. For example, Rhizobium melilotii will only nodulate alfalfa, while Rhizobium leguminosarum biovar trifolii will only nodulate clover (Trifolium). This host specificity is referred to cross inoculation group cell signaling between the bacteria and the legume host. The aforementioned Nod factors have been identified as lipochition oligosaccharides. Variations in the structures of these oligosaccharides determine the host specificity for the bacterium. Major cross inoculation groups are listed in Table 1.
Another example of the intricate relationship between the rhizobia and the host legume is the production of leghemoglobin (Appleby 1984). This molecule is similar in structure and function to hemoglobin found in human blood. It is only produced in fully functioning root nodules, and functions to bind and regulate the levels of oxygen in the nodule. Because the enzyme nitrogenase is sensitive to oxygen, free oxygen in nodule cell cytoplasm would inhibit the levels of nitrogen fixation in the nodule. Leghemoglobin seems to transport enough oxygen to allow the rhizobia to carry out cellular respiration, but not too much to inhibit the action of nitrogenase. This heme protein is jointly produced by the legume and bacterium; the legume produces the apoprotein while the bacterium produces the heme (porphyrin ring bound to an iron atom).
Figure 4
Extensive nodulation of a peanut root after inoculation with Bradyrhizobium strain 32H1.
Figure 5
Mutant non-nodulated soybeans (foreground) with normal, nodulated soybeans (background).
Figure 6
Comparison of peanut plants with and without Bradyrhizobia. Plants are (left to right), uninoculated with Bradyrhizobium, inoculated with Bradyrhibium, non-nodulating mutant peanut inoculated with Bradyrhizobium, and non-nodulating mutant peanut uninoculated with Bradyrhizobium.
The nodulation process illustrates an orchestrated interaction between the bacteria and host plant (Napoli & Hubbell 1975, Kamst et al. van Rhyn & Vanderleyden 1995, Cheng & Walker 1998). The process begins when the rhizobia are attracted to flavonoids released by the host legume’s roots. For legumes like alfalfa, clover, and soybeans (others like lupines and peanuts form nodules in other ways) the bacteria then begin to attach themselves to extensions of root epidermal cells called root hairs. The attachment process is actually a two-step process where the bacteria first attach using a Ca2+ - binding protein called rhicadhesin. After the bacteria accumulate and anchor themselves to the root hair surface, a firmer attachment that involves lectins and/or cellulose firbrils and fimbriae produced by the host plant and bacteria, respectively.
The host legume then senses chemicals produced by the rhizobia called Nod factors that cause the colonized root hairs to curl and form what is called a shepherd’s crook. Then rhizobia penetrate the root hairs and typically form a tubular structure called an infection thread. Once the bacteria reach the root itself, they stimulate cortical cell divisions that lead to the formation of a nodule. As the nodule begins to form, the bacteria become surrounded by a plant-derived membrane and are released inside plant cells forming the nodule. The bacteria subsequently lose their cell walls and undergo a profound change in cell morphology to form large, irregularly shaped branching cells called bacteroids. They then are entirely dependent on the host plant for their energy needs. In return, the bacteria fix nitrogen for the plant.
The interaction between the bacteria and host legume is so intricate that a particular Rhizobium or Bradyrhizobium will only nodulate a select number of plant genera. For example, Rhizobium melilotii will only nodulate alfalfa, while Rhizobium leguminosarum biovar trifolii will only nodulate clover (Trifolium). This host specificity is referred to cross inoculation group cell signaling between the bacteria and the legume host. The aforementioned Nod factors have been identified as lipochition oligosaccharides. Variations in the structures of these oligosaccharides determine the host specificity for the bacterium. Major cross inoculation groups are listed in Table 1.
| Crop | Nodulating Bacteria |
| Alfalfa | Sinorhizobium melilotii |
| Beans | Rhizobium legumninosarum biovar phaseoli and Rhizobium tropici |
| Clover | Rhizobium leguminosarum biovar trifolii |
| Lotus | Mesorhizobium loti |
| Peas | Rhizobium leguminosarum biovar viceae |
| Soybean | Bradyrhizobium japonicum, Bradyrhizobium elkanii, Rhizobium fredii |
| Sesbania | Azorhizobium caulinodans |
| Table 1. Major cross inoculation groups of legumes. | |
Another example of the intricate relationship between the rhizobia and the host legume is the production of leghemoglobin (Appleby 1984). This molecule is similar in structure and function to hemoglobin found in human blood. It is only produced in fully functioning root nodules, and functions to bind and regulate the levels of oxygen in the nodule. Because the enzyme nitrogenase is sensitive to oxygen, free oxygen in nodule cell cytoplasm would inhibit the levels of nitrogen fixation in the nodule. Leghemoglobin seems to transport enough oxygen to allow the rhizobia to carry out cellular respiration, but not too much to inhibit the action of nitrogenase. This heme protein is jointly produced by the legume and bacterium; the legume produces the apoprotein while the bacterium produces the heme (porphyrin ring bound to an iron atom).
Summary
Nitrogen is an essential nutrient for plant growth and development but is unavailable in its most prevalent form as atmospheric nitrogen. Plants instead depend upon combined, or fixed, forms of nitrogen, such as ammonia and nitrate. Much of this nitrogen is provided to cropping systems in the form of industrially produced nitrogen fertilizers. Use of these fertilizers has led to worldwide, ecological problems, such as the formation of coastal dead zones. Biological nitrogen fixation, on the other hand, offers a natural means of providing nitrogen for plants. It is a critical component of many aquatic, as well as terrestrial ecosystems across our biosphere.
References and Recommended Reading
Appleby, C. A. Leghemoglobin and Rhizobium respiration. Annual Review of Plant Physiology 33, 443-478 (1984).
Beijerinck, M. W. Über oligonitrophile Mikroben. Zbl. Backt. 7, 561-582 (1901).
Benson, D. R. & Silvester, W. B. Biology of Frankia strains, actinomycete symbionts of actinorhizal plants. Microbiological Reviews 57, 293-319 (1993).
Cheng, H-P. & Walker, G. C. Succinoglycan is required for initiation and elongation of infection threads during nodulation of alfalfa by Rhizobium meliloti. Journal of Bacteriology 180, 5183-5191 (1998).
Diaz, R. J. & Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 321, 926-929 (2008).
Fattah, Q. A."Plant Resources for Human Development." Third International Botanical Conference 2005. Bangladesh Botanical Society, Dhaka, Bangladesh, 2005.
Kamst, E. H., Spaink, P., & Kafetzopoulos, D. "Biosysnthesis and Secreation of Rhizobial Lipochitin-Oligosaccharide Signal Molecules," in Subcellular Biochemistry, Volume 29. Plant-Microbe Interactions, eds. B. B. Biswas & H. K. Das . (Plenum Publishing, New York, 1998) 29-33.
Herridge, D. F., Peoples, M. B., & Boddey, R. M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 311, 1-18 (2008).
Hubbell, D. H. & Kidder, G. Biological Nitrogen Fixation. University of Florida IFAS Extension Publication SL16. 1-4 (2009).
Napoli, C. A. & Hubbell, D. H. Ultrastructure of Rhizobium-induced infection threads in clover root hairs. Applied and Environmental Microbiology 30, 1003-1009 (1975).
National Research Council. Biological Nitrogen Fixation: Research Challenges.Washington, DC: National Academy Press, 1994.
Postgate, J. R. The Fundamentals of Nitrogen Fixation. New York, NY: Cambridge University Press, 1982.
Stephan, M. P. et al. "Physiological studies with Azospirillum spp. in Associative N2-Fixation. Volume I, eds. P. B. Vose & A. P. Ruschel (Boca Raton, FL: CRC Press, 1979) 7-14.
Vance, C. Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resouces. Plant Physiology 127, 391-397 (2001).
Van Rhyn, P. & Vanderleyden, J. The Rhizobium-plant symbiosis. Microbiological Reviews 59, 124-142 (1995).
Vadakattu, G. & Paterson, J. Free-living bacteria lift soil nitrogen supply. Farming Ahead 169, 40 (2006).
Vitousek, P. M. et al. Human alteration of the global nitrogen cycle: sources and consequences. Ecological Applications 7, 737-750 (1997). doi:10.1890/1051-0761(1997)007[0737:HAOTGN]2.0.CO;2
Vlassak, K. & Reynders, R. "Agronomic aspects of biological dinitrogen fixation by Azoxpirillum spp." in Associative N2 Fixation. Volume I, eds. P. B. Vose & A. P. Ruschel. (Boca Raton, FL: CRC Press, 1979) 93-102.










