
Abstract
Study design:
This is an experimental study.
Objectives:
The objective of this study was to evaluate the neuroprotective effects of Ginkgo biloba extract 761 (EGb761) on histological features of injured sites and on functional performance of rats subjected to standardized spinal cord injury (SCI).
Setting:
This study was conducted in Xian, Shaanxi, China.
Methods:
Thirty female Sprague–Dawley rats were randomly divided into three groups: sham-operated, saline-treated control and EGb761-treated. The Basso, Beattie, Bresnahan Locomotor Rating Score (BBB score) was calculated and footprint analysis was performed to evaluate the functional performance of the rats in each group. Hematoxylin and eosin (HE) staining and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) and caspase-3 staining were performed to evaluate the necrosis area and apoptotic cells at the injured site in each group.
Results:
At 14, but not 1, 3 and 7, days post injury (DPI), rats in the EGb761-treated group exhibited significantly better BBB scores compared with the saline-treated control group (P<0.05). The EGb761-treated group also showed increased stride length, decreased stride width and reduced toe dragging at 14 DPI (P<0.05). Analysis of HE staining revealed that the EGb761-treated group had reduced necrosis at the injury site compared with the saline-treated control group (P<0.05). Analysis of TUNEL and caspase-3 staining demonstrated that cell apoptosis was increased at 1–14 DPI, peaking at 24-h post injury in the gray matter, and 7 DPI in the white matter. At 7 DPI, the quantity of apoptotic cells was significantly decreased in the EGb761-treated group.
Conclusion:
EGb761 administration during the acute phase after SCI significantly reduced secondary injury-induced tissue necrosis and cell apoptosis and improved functional performance in rats.
Similar content being viewed by others


Introduction
Traumatic spinal cord injury (SCI) is a devastating condition that leads to severe neurological damage and requires extremely expensive, long-term care.1 SCI primarily affects young males with an annual incidence of 15–40 cases per million, causing a heavy burden for both family and society. The mechanism of SCI includes a primary injury and a subsequent secondary injury, which may have a long duration and may cause continuous damage to the function and structure of the spinal cord surrounding the primary injury region.2 Therefore, great effort has been made to explore treatment strategies for inhibiting the progression of secondary injury and protecting the remaining structure and function of the tissue surrounding the injured site. Even small anatomical gains can produce disproportionate functional benefits. For example, <10% of functional long-tract connections are necessary for locomotion.2, 3 Therefore, developing novel effective and convenient treatments for SCI is urgently required.
Ginkgo biloba extract 761 (EGb761) is a well-defined extract obtained from Ginkgo biloba leaves according to a standardized method. It has been used extensively for the treatment of diseases related to the central nervous system, including brain injury, neurodegenerative disorders and degenerative dementia.4, 5, 6 For example, EGb761 has neuroprotective effects and enhances neurogenesis after ischemic stroke.7, 8, 9 As an antioxidant agent, EGb761 not only has neuritogenic and angiogenic potential but also has the potential to augment repair and has regeneration mechanisms following stroke. However, the therapeutic effect of EGb761 on SCI has not been evaluated. It has been reported that secondary injuries following the initial impact include spinal cord ischemia, free-radical damage, apoptosis, electrolyte imbalance, excitotoxicity and inflammatory injury.10, 11, 12 Given the antioxidant activity of EGb761, and the potential of EGb761 to promote regeneration after stroke, it is theoretically and practically important to investigate whether EGb761 has beneficial effects for SCI. Here, we systematically evaluated the effects of EGb761 on histological and functional recovery in rats after SCI.
Materials and methods
Animals
Thirty female Sprague–Dawley rats (200–220 g, provided by the Animal Center of the Fourth Military Medical University) were randomly divided into three groups—(1) sham-operated: rats were subjected to laminectomy but not SCI injury control group; (2) saline-treated control: SCI rats receiving saline intraperitoneally (i.p.); (3) EGb761-treated: SCI rats receiving EGb761 (Chi Sheng Chemical Corporation, Taiwan, China) at a dose of 100 mg kg−1 day−1 i.p. All of the rats were maintained on a 12–12 h light–dark cycle and were allowed free access to food and water. The procedures were reviewed and approved by the Animal Care Committee of the Fourth Military Medical University. All efforts were made to minimize the number of animals used and their suffering. At the end of the studies, all rats were killed with overdose of sodium pentobarbital.
Surgical procedure
Surgical procedure was performed as described previously.13 The rats were anesthetized with 1% sodium pentobarbital (50 mg kg–1) i.p. A 30–40 mm dorsal midline incision was made at the T8–T12 level, and bilateral laminectomy was performed to expose the vertebrate T10 spinal cord. The NYU Impactor (MASCIS, New York University, New York, NY, USA) was used to produce a contusion SCI and to monitor the contusion.13 After being placed in the Impactor frame, the T8 and T11 processes were clamped, and the Impactor rod was adjusted to the zero position, after which the basal clip was attached to the border of the surgical wound. The Impactor was then lowered between the upper margin of T9 and the lower margin of T10. After contacting with the spinal cord, as indicated by means of audible and visible signals of the device caused by closing the circuit between the basal clip and the Impactor head, the Impactor rod was raised to a height of 25 mm. The height and date information were entered into the Impactor program. The Impactor rod was then released to hit the spinal cord, which was lifted off immediately after producing the contusion. The contusion site was inspected, active bleeding was stopped and the paravertebral muscles and the skin were closed. Then, the operated animals were kept in cages with soft bedding and ambient temperature maintained at 20–23 degrees.
Drug administration
An hour after the injury, the rats received i.p. administration of saline (control group), at a dose of 100 mg kg–1 bodyweight per day of EGb761. Over the days following the injury, the rats received daily doses of the same vitamins, until the day they were killed. All rats received postoperative treatment including the i.p. injection of penicillin for a week. Bladders were manually voided three times daily until reflex bladder emptying was established. The rats were maintained for 4 weeks under postoperative care recommended by the NIH Guide for Care and Use of Laboratory Animals and were given access to food and water ad libitum.
Functional assessment
The BBB Locomotor Rating Scale was used to evaluate the recovery of hindlimb locomotor functions in an open field,14 as reported previously.13 Briefly, the behavioral evaluations were carried out at 0800–0900 hours weekly for 4 weeks after operation. The BBB locomotor rating scale ranges from 0, which corresponds to flaccid paralysis, to 21, which is normal gait. Rats were allowed to walk around freely in a spacious field for 5 min when the movement of the hindlimbs was closely observed. Three observers blind to the animals’ identity assisted in this task. The average of the three scores of the three observers was calculated. Before testing, bladders were expressed as spontaneous bladder contraction often accompanied by hindlimb activity.
Footprint analysis
The analysis was conducted weekly for 4 weeks after operation, as reported previously.13 Briefly, the plantar surface of both hindlimbs of each rat was colored black and the dorsal surface was colored red with nontoxic inks. The rat was then allowed to run toward a dark tunnel on the white paper (21 × 59.4 cm2), and thus the red and/or black ink was printed onto the paper as one set of footprints.2 From each set of footprints, the pixels of the black (normal plantar paw placing) and red (abnormal dorsal paw placing) areas were determined using ImageJ (http://rsb.info.nih.gov/ij/). The percentage of red pixels defined as red/(black+red) × 100% was computed to quantify toe dragging. Stride length (distance between the centers of ipsilateral adjacent footprints) and stride width (perpendicular distance between the centers of left and right hind limbs) were measured, and the average of five steps in each case was used for statistical comparison.
Histological investigation of the spinal cord
At 4 weeks after SCI, the rats were anesthetized with i.p. injection of 1% sodium pentobarbital (50 mg kg–1) as described above, and 1 liter of 0.1 mmol l–1 phosphate-buffered saline was introduced into the left ventricle for 20–30 min, followed by 1 liter of 4% paraformaldehyde in 0.1 mol l–1 phosphate-buffered saline (pH 7.2–7.3, 4 oC for 1 h). The spinal cords were taken out and immersed in the same fixative at 4 oC for 24 h, and then transferred to phosphate buffer containing 30% sucrose before cryosectioning. Serial sections of the spinal cord tissues were cut at a thickness of 20 μm (some sagitally and others coronally) in a cryo-microtome (Leica CM 1900, Leica Microsystems Nussloch GmbH, Nussloch, Germany) and mounted on gelatin-pretreated slides for the following laboratory procedures. Hematoxylin and eosin (HE) staining was performed for histological analysis.
Quantification of the area of injury
HE-stained sections were used to evaluate the necrosis area in the injured site. Five sections were chosen for analysis in each rat, namely, the section with central canal and two adjacent sections on both sides, with 200 μm between each section. The microscopic images were obtained with a 40 × objective under an Olympus BX-51 microscopy (Olympus Corporation, Tokyo, Japan).
TUNEL examination
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) examination was performed according to the manufacturer’s protocol (Roche Company, Grenzacherstrasse, Switzerland). A TUNEL reaction mixture was made and 50 μl was added to each slide. Tow positive control and two negative controls were performed for each staining. For positive controls, DNA fragmentation was induced by incubation with DNase I for 10 min at room temperature before staining. For negative controls, solutions without terminal deoxynucleotidyl transferase was added for reaction. The slides were investigated under a fluorescence microscope, and images were captured for quantitative analysis. Five slides were used for TUNEL staining for each animal. Eight high-power visual fields (× 200) were selected for each slide (four in the gray matter, and the other four in the white matter) to calculate the apoptotic index (the number of TUNEL-positive cells divided by the total number of cells).
Immunohistochemistry analysis
Serial tissue sections were immunostained using rabbit monoclonal antibody against caspase-3 (Cell Signaling, Danvers, MA, USA). The sections were blocked for endogenous peroxidase and biotin before the application of the primary antibody. Incubation of primary antibodies was performed overnight at 4 oC. The Elite Vector Stain ABC System (Vector Laboratories, Burlingame, CA, USA) was used as the detection system and diaminobenzidine was used as a chromogen. The nucleus was counterstained with hematoxylin for quantification. For each section, eight high-power visual fields (× 400) were selected (four in gray matter, and the other four in white matter) to calculate the percentage of caspase-3-positive cells (the number of caspase-3-positive cells divided by the total number of cells).
Statistical analysis
All data were expressed as mean±s.d. Statistical analyses were performed using Student’s t-tests and one-way analysis of variance with least squared difference post-hoc tests, as appropriate. All P-values are two-tailed. A value of P<0.05 was considered statistically significant. Statistical analyses were performed using SPSS v.13.0.0 (SPSS, Inc., Chicago, IL, USA).
Results
BBB scoring
The functions of the posterior limbs of all rats were normal before SCI, with a BBB score of 21. The rats in the sham-operated group showed slight somatic instability at 1–3 days post injury (DPI), which was probably caused by operational pain and stimulation. The rats had completely recovered by 7 DPI, indicating that laminectomy alone does not affect the BBB score of the rats. The motor function of rats in both the saline-treated control group and EGb761-treated group was completely lost after SCI, with a first BBB score of 0. The functions of the posterior limbs of rats in these two groups began to recover after 3 DPI. At 7 DPI, the hip, knee and ankle joints of the rats in these two groups showed substantial activity. At this time point, the BBB score for the EGb761-treated group was higher than that of the saline-treated control (7.60±1.14 vs 7.80±0.84; Figure 1), although this difference was not statistically significant. At 14 DPI, the BBB scores for the EGb761-treated group were significantly higher than those of the saline-treated control group (15.40±0.89 vs 13.60±0.55, P<0.05; Figure 1).

Graph showing the BBB score of each group at different time points after SCI. *P<0.05.
Footprint analysis
Toe dragging, stride length and stride width were further analyzed to evaluate the functional recovery of rats in each group at 7 and 14 days after operation (Figure 2). As shown in Figure 2, sham-operated rats showed consistent plantar stepping during the entire process (Figure 2a), whereas severe toe dragging with no planar stepping was observed in the control group at 14 DPI (Figure 2b). After SCI, the stride length increased and the stride width decreased progressively. However, the EGb761-treated group exhibited greater increased stride length, decreased stride width and less toe dragging compared with the saline-treated control group (Figures 2, 3 and 4).

Representative images showing the footprint for each group at 14 days after SCI. (a) Sham-operated rat; (b) saline-treated control group (SCI+SAL); (c) EGb761-treated group (SCI+EGb761). A full color version of this figure is available at the Spinal Cord journal online.

Graph showing the quantitative results of footprint analysis (stride length and stride width) for each group at 7 (a) and 14 (b) DPI. *P<0.05.

Graph showing the quantitative results of toe dragging ratio of each group at different time points after SCI. *P<0.05.
At 7 DPI, stride length and stride width were not significantly different between the EGb761-treated group and the saline-treated control group (Figure 3), whereas the ratio of rats with toe dragging in the EGb761-treated group was lower than that for the control group, although this difference was not statistically significant (93.10±1.22 vs 95.68±2.17, P>0.05; Figure 4). Noticeably, at 14 DPI, compared with the saline-treated control group, the EGb761-treated group exhibited increased stride length (10.44±0.26 vs 9.81±0.33, P<0.05; Figure 3), decreased stride width (4.52±0.14 vs 5.58±0.11, P<0.05; Figure 3) and decreased ratio of rats with toe dragging (50.86±3.27 vs 62.40±3.24, P<0.05; Figure 4).
Histological outcome measures
HE-staining analysis
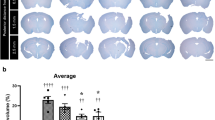
The rats were killed at 7 or 14 days after SCI, and the spinal cords were sagittally sectioned and stained with HE (Figure 5). At 14 DPI, extensive necrosis and prominent cavitation were observed at the injury site in the control group (Figure 5a), whereas the necrosis and cavitation areas were much smaller in the EGb761-treated group (Figure 5b). Quantitative analysis revealed that the lesion area (including the necrosis and cavitation area) in the EGb761-treated group was much smaller than that of the control group (2.92±0.18 vs 3.63±0.27 cm2, P<0.05; Figure 5c).

Representative images and quantitative results showing the lesion area at the section with central canal in each group at 14 DPI. The lesion area was outlined with dashed lines. (a) Saline-treated control group; (b) EGb761-treated group; (c) the quantitative results of the lesion area in each group. *P<0.05. A full color version of this figure is available at the Spinal Cord journal online.
Apopotosis analysis
TUNEL staining and caspase-3 staining were performed to examine the apoptotic cells after SCI. Very few, if any, TUNEL-positive apoptotic cells were observed in the sham-operated group. For the rats in the control group, the number of TUNEL-positive apoptotic cells in both the gray matter and white matter around the injured site were increased after SCI, exhibiting special quantity patterns at different time points (Figure 6,Table 1). The apoptosis of cells in the gray matter primarily occurred during the first 24 h, whereas apoptosis of cells in the white matter began to increase at 3 DPI, and peaked at 7 DPI (Figure 6,Table 1). The trend of cell apoptosis in the EGb761-treated group was similar, except that the quantity of apoptotic cells was significantly reduced at 3 and 7 DPI compared with the control group (3d: 27.84±3.01 vs 33.86±2.33, P<0.05; 7d: 58.12±2.96 vs 69.52±4.74, P<0.05; Figures 7 and 9). Analysis of caspase-3-positive apoptotic cells revealed similar results (Figures 8 and 9).

Number of TUNEL-positive apoptotic cells in gray matter and white matter at different time points after SCI. *P<0.05.
 , n=5)
, n=5)
Representative images of TUNEL-positive cells in gray matter and white matter at different time points after operation in sham-operated (a), saline-treated (b1, b2) or EGb761-treated (c1, c2) groups (× 200). A full color version of this figure is available at the Spinal Cord journal online.

Quantitative results of caspase-3 (a) and TUNEL-positive (b) cells at different time points after SCI in saline-treated or EGb761-treated groups.

Representative images of caspase-3-positive cells at different time points after SCI in saline-treated (a1, a2) or EGb761-treated (b1, b2) groups (× 400). A full color version of this figure is available at the Spinal Cord journal online.
Discussion
This study investigated the therapeutic effect of EGb761 administration on an acute SCI rat model, and found that it improves histological and functional recovery after SCI.
Apoptosis of cells is an important process during secondary injury after SCI, leading to continuous neuron cell loss and degeneration, which in turn deteriorates the injury.14 Here, we demonstrated that cell apoptosis after SCI exhibited special temporal and spatial features. Apoptosis in the gray matter around the injury site peaked at 1 DPI, whereas apoptosis in the white matter began to increase and spread to the surrounding tissue at 1 DPI, and peaked at 7 DPI. White matter contains mainly microglia and oligodendrocytes, which are closely related to the myelination of axons. Therefore, loss of oligodendrocytes accelerates the degeneration of axons.15
It has been reported that EGb761 has anti-apoptosis potential in neurodegenerative diseases through different kinds of mechanisms.16 The anti-apoptosis activity of EGb761 is primarily induced by the constituents flavonoid and terpenoid. In addition, EGb761 has beneficial neuroprotective effects, probably by means of its antioxidant action, which involves the scavenging of free radicals.10, 11, 12, 17 In this study, we found that the number of apoptotic cells at 3 and 7 DPI in the EGb761-treated group was significantly smaller than that in the control group. HE-staining analysis further revealed that the cavity in the EGb761-treated group was significantly reduced. Therefore, EGb761 treatment reduced the number of apoptotic cells in the white matter around the injury site and improved tissue recovery after SCI.
Caspase-3 is a common downstream effector of different apoptosis pathways, and it is an indispensable part of the apoptosis caspase cascade. Caspase-3 is synthesized and stored in the form of proenzyme, and it is activated during the reaction of the apoptosis caspase cascade. Therefore, caspase-3 is often used as an early biochemical index.18 To reduce apoptosis after SCI, therapeutic interventions should be given in the critical time window between the time of injury and the activation of caspase-3. Here, we demonstrated that the temporal and spatial distribution of caspase-3-positive cells is consistent with that of TUNEL apoptotic cells, whereas EGb761 treatment in the early stage after SCI significantly reduced the number of caspase-3-positive apoptotic cells. Therefore, in the acute rat SCI model in this study, EGb761 treatment exerted neuroprotective effects by at least partially inhibiting cell apoptosis by reducing the expression of caspase-3. Nevertheless, it should be noted that the mechanism of apoptosis is a complex process involving a large number of cytokines and signal transduction molecules, which is not fully understood. Therefore, the mechanisms of EGb761-induced inhibition of apoptosis are not completely clear, and further studies are needed to explore whether EGb761-induced inhibition of caspase-3 is related to its anti-apoptosis activity.
After SCI, the number of neurons reduces and the necrosis cavity enlarges gradually. However, the motor function of the posterior limb improves. The mechanisms behind this phenomenon are suggested to be related to the remaining axons and axonal regeneration after SCI.19 In this study, we observed that at 14 DPI, the morphology of tissue around the injury site recovered to a certain degree, as indicated by the increased number of glial cells and reduction of vacuolar degeneration of the myelin sheath, implying that the functional recovery of the posterior limb might be related to the remyelination and regeneration of the remaining axons. The functional assays revealed that there was no difference between the sham-operated rats and normal rats, indicating that laminectomy alone does not result in functional defects. By comparing the functional results from different groups, we found that the EGb761-treated group exhibited better results for stride length, stride width and ratio of toe dragging compared with saline-treated control group. In addition, at 14 DPI, the necrosis cavity areas in the EGb761-treated group were significantly smaller than those of the saline-treated control group. Therefore, EGb761 can inhibit cell apoptosis in the early stage after SCI, reducing necrosis cavity area and ultimately improving neurological function recovery.
In conclusion, EGb761 administration in the acute phase after SCI significantly reduced secondary damage-induced tissue necrosis and improved the behavioral function of the rats. These results suggest that EGb761 has neuroprotective effects after SCI, providing novel feasible therapeutic strategies for acute SCI treatment. Nevertheless, some issues should be further explored in future studies before clinical administration of EGb761 after SCI. For example, the optimal time period after SCI for treatment with EGb761 should be determined to obtain better therapeutic results.
Data archiving
There were no data to deposit.
References
Thuret S, Moon LD, Gage FH . Therapeutic interventions after spinal cord injury. Nat Rev Neurosci 2006; 7: 628–643.
Saadoun S, Bell BA, Verkman AS, Papadopoulos MC . Greatly improved neurological outcome after spinal cord compression injury in AQP4-deficient mice. Brain 2008; 131: 1087–1098.
Blight AR . Cellular morphology of chronic spinal cord injury in the cat: analysis of myelinated axons by line-sampling. Neuroscience 1983; 10: 521–543.
Montes P, Ruiz-Sanchez E, Rojas C, Rojas P . Ginkgo biloba extract 761: a review of basic studies and potential clinical use in psychiatric disorders. CNS Neurol Disorders Drug Targets 2015; 14: 132–149.
Shi C, Xiao S, Liu J, Guo K, Wu F, Yew DT et al. Ginkgo biloba extract EGb761 protects against aging-associated mitochondrial dysfunction in platelets and hippocampi of SAMP8 mice. Platelets 2010; 21: 373–379.
Cho JH, Sung JH, Cho EH, Won CK, Lee HJ, Kim MO et al. Gingko biloba extract (EGb 761) prevents ischemic brain injury by activation of the Akt signaling pathway. Am J Chin Med 2009; 37: 547–555.
Nada SE, Tulsulkar J, Shah ZA . Heme oxygenase 1-mediated neurogenesis is enhanced by Ginkgo biloba (EGb 761(R)) after permanent ischemic stroke in mice. Mol Neurobiol 2014; 49: 945–956.
Nada SE, Shah ZA . Preconditioning with Ginkgo biloba (EGb 761(R)) provides neuroprotection through HO1 and CRMP2. Neurobiol Dis 2012; 46: 180–189.
Zhang Z, Peng D, Zhu H, Wang X . Experimental evidence of Ginkgo biloba extract EGB as a neuroprotective agent in ischemia stroke rats. Brain Res Bull 2012; 87: 193–198.
Jia Z, Zhu H, Li J, Wang X, Misra H, Li Y . Oxidative stress in spinal cord injury and antioxidant-based intervention. Spinal Cord 2012; 50: 264–274.
Cristante AF, Barros Filho TE, Oliveira RP, Marcon RM, Rocha ID, Hanania FR et al. Antioxidative therapy in contusion spinal cord injury. Spinal Cord 2009; 47: 458–463.
Hall ED . Antioxidant therapies for acute spinal cord injury. Neurotherapeutics 2011; 8: 152–167.
Yan M, Yang M, Shao W, Mao XG, Yuan B, Chen YF et al. High-dose ascorbic acid administration improves functional recovery in rats with spinal cord contusion injury. Spinal Cord 2014; 52: 803–808.
Zhang N, Yin Y, Xu SJ, Wu YP, Chen WS . Inflammation & apoptosis in spinal cord injury. Indian J Med Res 2012; 135: 287–296.
Mekhail M, Almazan G, Tabrizian M . Oligodendrocyte-protection and remyelination post-spinal cord injuries: a review. Prog Neurobiol 2012; 96: 322–339.
Sun L, Zhuang W, Xu X, Yang J, Teng J, Zhang F . The effect of injection of EGb 761 into the lateral ventricle on hippocampal cell apoptosis and stem cell stimulation in situ of the ischemic/reperfusion rat model. Neurosci Lett 2013; 555: 123–128.
Jiang X, Nie B, Fu S, Hu J, Yin L, Lin L et al. EGb761 protects hydrogen peroxide-induced death of spinal cord neurons through inhibition of intracellular ROS production and modulation of apoptotic regulating genes. J Mol Neurosci 2009; 38: 103–113.
Choudhary GS, Al-Harbi S, Almasan A . Caspase-3 activation is a critical determinant of genotoxic stress-induced apoptosis. Methods Mol Biol 2015; 1219: 1–9.
Beattie MS, Bresnahan JC, Komon J, Tovar CA, Van Meter M, Anderson DK et al. Endogenous repair after spinal cord contusion injuries in the rat. Exp Neurol 1997; 148: 453–463.
Acknowledgements
This study was partially supported by the National Natural Science Foundation of China (81301581).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Yan, M., Liu, Yw., Shao, W. et al. EGb761 improves histological and functional recovery in rats with acute spinal cord contusion injury. Spinal Cord 54, 259–265 (2016). https://doi.org/10.1038/sc.2015.156
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sc.2015.156
