
Abstract
Orexins (OX) and their receptors (OXR) modulate feeding, arousal, stress, and drug abuse. Neural systems that motivate and reinforce drug abuse may also underlie compulsive food seeking and intake. Therefore, the effects of GSK1059865 (5-bromo-N-[(2S,5S)-1-(3-fluoro-2-methoxybenzoyl)-5-methylpiperidin-2-yl]methyl-pyridin-2-amine), a selective OX1R antagonist, JNJ-10397049 (N-(2,4-dibromophenyl)-N′-[(4S,5S)-2,2-dimethyl-4-phenyl-1,3-dioxan-5-yl]urea), a selective OX2R antagonist, and SB-649868 (N-[((2S)-1-{[5-(4-fluorophenyl)-2-methyl-1,3-thiazol-4-yl]carbonyl}-2-piperidinyl)methyl]-1-benzofuran-4-carboxamide), a dual OX1/OX2R antagonist were evaluated in a binge eating (BE) model in female rats. BE of highly palatable food (HPF) was evoked by three cycles of food restriction followed by stress, elicited by exposing rats to HPF, but preventing them from having access to it for 15 min. Pharmacokinetic assessments of all compounds were obtained under the same experimental conditions used for the behavioral experiments. Topiramate was used as the reference compound as it selectively blocks BE in rats and humans. Dose-related thresholds for sleep-inducing effects of the OXR antagonists were measured using polysomnography in parallel experiments. SB-649868 and GSK1059865, but not JNJ-10397049, selectively reduced BE for HPF without affecting standard food pellet intake, at doses that did not induce sleep. These results indicate, for the first time, a major role of OX1R mechanisms in BE, suggesting that selective antagonism at OX1R could represent a novel pharmacological treatment for BE and possibly other eating disorders with a compulsive component.
Similar content being viewed by others
INTRODUCTION
Episodes of binge eating (BE) in humans are characterized by compulsive, non-homeostatic consumption of an unusually large quantity of highly palatable food (HPF) in a short period of time. Even though they are not hungry, subjects eat more rapidly than normal until feeling uncomfortably full. As described by the DMS-IV-TR (American Psychiatric Association, 2000), these episodes are accompanied by a subjective sense of loss of control over eating, and are associated with feeling of distress, disgust, depression, being guilty about overeating, and eating alone because of embarrassment.
BE represents a central feature of bulimia nervosa, in which episodes of BE are followed by behaviors aimed at avoiding weight gain, such as self-induced vomiting. Intense and persistent BE episodes represent typical phenomena also occurring in subjects suffering from binge-eating disorder (BED) (Walsh and Devlin, 1998). BED is characterized by repeated episodes of BE in the absence of compensatory behaviors to avoid weight gain. The diagnostic criteria for BED in the DSM-IV-TR indicate that BE episodes should occur at least 2 days per week for 6 months. BED is associated with significant medical and psychiatric co-morbidity (Javaras et al, 2008; Grucza et al, 2007; Fassino et al, 2003). It is estimated that BE afflicts approximately 5% of the US adult population at some time during their lives (Foulds Mathes et al, 2009) and it contributes to aggravate obesity and associated pathologies (Hudson et al, 2007; Heath, 1998; Devlin et al, 2000; Yanovski, 2003).
Current medications, like topiramate (McElroy et al, 2007; McElroy et al, 2009) or sibutramine (Appolinario et al, 2000; Wilfley et al, 2008), have been reported to reduce BE in clinical studies. However, their administration is associated with a variety of adverse side effects, which represent serious problems during chronic treatment (McElroy et al, 2009; Carter et al, 2003; Yager, 2008). In particular, sibutramine has been recently withdrawn from the European market, while topiramate is well known for its cognitive impairing properties. Innovative treatments for bulimia nervosa and BED without severe side effects are strongly needed.
In 1998, two groups independently identified a new class of neuropeptides originating in hypothalamic nuclei (Sakurai et al, 1998; de Lecea et al, 1998). These peptides, called orexin-A (OXA) and orexin-B (OXB) (also denoted a hypocretin 1 and hypocretin 2), are produced from proteolytic processing of the pre-pro-OX peptide and bind to two GPCRs, namely OX-1 and OX-2 receptors (OX1R and OX2R) (also denoted as HcrtR1 and HcrtR2). OX1R is coupled to Gq/11, whereas studies using neuronal cells suggest that OX2R is coupled to Gq, Gs, and Gi proteins. In the central nervous system, OX1R and OX2R show partially overlapping, but largely distinct and complementary, distribution patterns (Sakurai, 2007). Brain areas such the infralimbic cortex, hippocampus, and locus coeruleus exhibit high expression of OX1R, whereas OX2R is the only receptor expressed in arcuate nucleus, tuberomammillary nucleus, and dorsomedial and lateral hypothalamus (LH). Both receptors are present in the prefrontal cortex, amygdala, bed nucleus of stria terminalis, paraventricular thalamic nucleus, dorsal raphe, ventral tegmental area (VTA), and laterodorsal tegmental nucleus–peduncolo pontine nucleus (Lu et al, 2000; Marcus et al, 2001; Trivedi et al, 1998). These findings suggest that OXs and their receptors are likely to play a broad regulatory role in the central nervous system.
The physiology of the awake/sleep state is one of the fields in which the role of OXs has been most extensively studied. Indeed, the disruption of OX signaling in prepro-OX knockout mice produced a phenotype with characteristics remarkably similar to those of patients suffering from narcolepsy, a chronic disorder characterized by excessive sleepiness that may be associated with very severe sleep paralysis, hypnagogic hallucinations, and cataplexy (Chemelli et al, 1999). The excessive sleepiness appears to be a manifestation of an inability to maintain extended waking.
Moreover, the simultaneous antagonism of both OX1R and OX2R or the selective inhibition of OX2R results in the induction of a strong hypnotic effect (Brisbare-Roch et al, 2007; Dugovic et al, 2009; Di Fabio et al, 2011).
Data in the literature also support a role of the OX system in feeding behavior, in the control of both homeostatic and reward-based food intake. Further to exhibiting a narcolepsy phenotype, OX knockout mice are also hypophagic compared with weight and age-matched littermates, indicating a role for OXs in modulating feeding and energy metabolism (Willie et al, 2001). Injection of OXA into the lateral ventricle of rats, during early light phase, induced a dose-related increase of food intake in rats (Sakurai et al, 1998), which was blocked by pre-treatment with the OX1R antagonist SB-334867 (Haynes et al, 2000; Rodgers et al, 2001). The role of OXs in reward-based food intake has been documented by a recent paper of Perello et al (2010), showing that the increase in the rewarding value of high-fat diet induced by ghrelin is OX-dependent; moreover, SB-334867 has been reported to inhibit high-fat food self-administration (Nair et al, 2008). Activation of the OX1R is a necessary component of food-reinforced responding, motivation, or both (Sharf et al, 2010). In addition, LH OX neurons are activated by cues associated with consummatory rewards such as food (Harris et al, 2005), suggesting a potential role of the OX system in response to external environmental cues linked to cognitive aspects of feeding.
Recent reports support a role for OX signaling in the neurobehavioral and motivational effects of drugs of abuse (Harris et al, 2005; Borgland et al, 2006; Jupp et al, 2011; for a review, see Bonci and Borgland, 2009; Martin-Fardon et al, 2010). Thus, blockade of OX1R decreases ethanol (Lawrence et al, 2006) and nicotine self-administration (Hollander et al, 2008), inhibits cue-induced reinstatement of ethanol- (Lawrence et al, 2006), cocaine- (Smith et al, 2010), and morphine-seeking (Harris et al, 2005), and attenuates stress-induced reinstatement of cocaine- (Boutrel et al, 2005) and ethanol-seeking (Richards et al, 2008). Moreover, recent evidence also linked OX2R-selective mechanisms to alcohol reward and seeking behavior (Shoblock et al, 2011).
Evidence is accumulating that excessive intake of certain foods under specific conditions produces behaviors and changes in the brain that resemble an addiction-like state (Gold et al, 2003; Kenny, 2011; Pelchat et al, 2004; Avena et al, 2008; Ifland et al, 2009; Gearhardt et al, 2011a). Neural systems that motivate and reinforce drug abuse have also been proposed to underlie behaviors associated with compulsive food seeking and food intake (Johnson and Kenny, 2010; Hoebel, 1985; Volkow and Wise, 2005; Corwin et al, 2011; Gearhardt et al, 2011b; Wang et al, 2011). These findings raise the question of whether the OX system may also have a role in eating disorders characterized by compulsive binge-type episodes, such as bulimia nervosa and BED.
Therefore, this study was aimed at investigating the effect of the dual OX1/OX2R antagonist SB-649868 (N-[((2S)-1-{[5-(4-fluorophenyl)-2-methyl-1,3-thiazol-4-yl]carbonyl}-2-piperidinyl)methyl]-1-benzofuran-4-carboxamide) (Di Fabio et al, 2011), the selective OX1R antagonist GSK1059865 (5-bromo-N-[(2S,5S)-1-(3-fluoro-2-methoxybenzoyl)-5-methylpiperidin-2-yl]methyl-pyridin-2-amine) (Gozzi et al, 2011), and the selective OX2R antagonist JNJ-10397049 (N-(2,4-dibromophenyl)-N′-[(4S,5S)-2,2-dimethyl-4-phenyl-1,3-dioxan-5-yl]urea) (McAtee et al, 2004; Dugovic et al, 2009) in the BE model described by Cifani et al (2009), in which BE episodes for HPF is evoked in female rats by cycles of food restriction/re-feeding and acute stress. The three antagonists were first evaluated in vitro in rat recombinant OX1R and OX2R to determine their potency and to confirm their selectivity for the two receptor subtypes. Their pharmacokinetics (PKs) were then evaluated and the doses able to induce hypnotic effects were determined in a sleep rat model. Finally, the compounds were tested at the defined doses in the BE model.
MATERIALS AND METHODS
Animals
All research involving animals were carried out in accordance with the European directive 86/609/EEC governing animal welfare and protection, which is acknowledged by the Italian Legislative Decree no. 116, 27 January 1992, and according to internal review performed by the GlaxoSmithKline Committee on Animal Research & Ethics (CARE) and to the company Policy on the Care and Use of Laboratory Animals.
Drugs
SB-649868 (Di Fabio et al, 2011), GSK1059865 (Gozzi et al, 2011), and JNJ-10397049 (McAtee et al, 2004) were synthesized in GSK laboratories. OXA was supplied by California Peptides Research (cat. no. 471-99, CA). Myo-[1,2-3H(N)]inositol (NET-906, specific activity: 51 Ci/mol) and yttrium silicate RNA binding beads (RPNQ0013) were purchased from Perkin-Elmer (Italy). Topiramate (Topamax; Janssen-Cilag) was purchased from Janssen-Cilag. It was available in tablets, which were reduced into powder before administration.
Experiment 1: Antagonism of SB-649868, JNJ-10397049, and GSK1059865 at Rat OX1R and Rat OX2R
Cell culture
Rat basophil leukemia cells stably transfected with rat OX1R (rOX1R) or rat OX2R (rOX2R) were cultured in α-MEM (Invitrogen/GIBCO) supplemented with 10% fetal bovine serum (FBS, PAA), 100 U/ml penicillin G, 100 U/ml streptomycin (Pen/Strep; Invitrogen/GIBCO), and 400 μg/ml geneticin (Invitrogen/GIBCO), at 37 °C with 5% CO2 in a humidified atmosphere.
Accumulation of [3H]inositol phosphates (IPs)
Accumulation of [3H]inositol phosphates (IPs) was measured as described previously (Brandish et al, 2003) with the following modifications. The cell lines stably expressing rOX1R or rOX2R were seeded in 96-well tissue culture plates at 3 × 104 cells per well and 1.5 × 104 cells per well, respectively, in α-MEM supplemented with 10% FBS and Pen/Strep without geneticin. After 24 h, culture medium was aspirated and 100 μl of fresh medium supplemented with 10 μCi/ml NET-906 (Perkin-Elmer) was added to cells; thus, 1 μCi of radiolabeled inositol was used per well. After 16 h of incubation, cells were washed two times with the assay buffer (1 × HBSS, 20 mM HEPES (pH 7.4), plus 0.1% bovine serum albumin and 10 mM LiCl), before the addition of agonists or antagonists. Antagonists were incubated for 30 min at 37 °C before stimulation with agonist. Concentration–response curves (CRCs) of OXA ranging from 0.0001 to 10 μM were performed. After 1 h of incubation at 37 °C, the assay buffer was aspirated, 80 μl per well of 0.1 M ice-cold formic acid were added, and cells were left for 30 min incubation at room temperature. A measure of 20 μl of the cell extract was added to 80 μl of yttrium silicate beads (YSi SPA; Perkin-Elmer; 12.5 mg/ml), shaken for 1 h at room temperature, and left at 4 °C for 2 h before counting on a Packard Top-Count NXT Microplate Scintillation Counter.
Data were expressed as % maximal agonist response calculated as follows: %maximal agonist response=((c.p.m.antagonist−c.p.m.basal)/c.p.m.maxagonistresponse−c.p.m.basal) × 100.
In-vitro data analysis
CRCs were fitted by sigmoidal nonlinear regression analysis using the GraphPad Prism 5.0 software (GraphPad Software, San Diego, CA) to obtain agonist EC50 (agonist concentration required to obtain 50% of the maximal response).
The potency (KB=antagonist dissociation constant) of non-surmountable antagonists was determined by applying the operational model equation for non-competitive antagonism (Kenakin et al, 2006). The KB value for surmountable antagonists was calculated by Schild's analysis (Arunlakshana and Schild, 1959). For SB-649868 only the antagonist IC50 was calculated. We plotted the response produced by 1 μM OXA in the absence and in the presence of four different concentrations of antagonist. IC50 is defined as the antagonist concentration required to inhibit by 50% the response produced by the agonist. Results are expressed as pEC50 (−log10 EC50), pKB (−log10 KB), or pIC50 (−log10 IC50), and they are presented as mean±SEM or as mean with 95% confidence limits (95% CL) of at least three independent experiments. All drugs tested were dissolved in dimethyl sulfoxide (DMSO) and further diluted in the assay buffer to give a final DMSO concentration not exceeding 0.5%.
Experiment 2: PK Determinations in Male and Female Rats
To evaluate PK blood exposure for the sleep and BE studies, PK profiles of compounds have been analyzed in male and female rats in the same experimental conditions of sleep and BE experiments. The PK profiles were investigated after administration by gavage of 3 mg/kg SB-649868 in female and male rats, intraperitoneal administration of 10 mg/kg in female and 5 mg/kg in male rats of JNJ-10397049, and administration by gavage in female rats and intraperitoneal administration in male rats of 10 mg/kg of GSK1059865. Blood samples were collected via the femoral vein at intervals up to 4 h after administration. Brain samples were collected at the end of the experiment. The concentration of SB-649868, JNJ-10397049, and GSK1059865 in blood and brain samples was determined using a method based on protein precipitation followed by HPLC-MS/MS analysis. Non-compartmental PK parameters were obtained from the blood concentration−time profiles using the software package WinNonlin v.4.0 (Pharsight, Mountain View, CA). PK parameters are expressed as mean±SD (Read and Braggio, 2010).
Experiment 3: Effect of SB-649868, JNJ-10397049, and GSK1059865 on Rat Sleep Model
Animals
Male Sprague–Dawley rats (275–300 g; Charles River, Calco, Como, Italy) were housed singly on a 12-h light–dark cycle (light on at 0300 h) 1 week before surgery. Access to food and water was allowed ad libitum. To collect the biopotential signals, a miniature multichannel telemetric transmitter (TL10M3-F40-EET; Data Sciences Int.) was implanted intraperitoneally into the animals. Two electrodes were fixed permanently with dental cement to the skull to record cortical electroencephalogram (EEG). They were directly in contact with the dura mater through two drilled holes on the fronto-parietal region. Two other electrodes were fixed to the skeletal muscles of the neck, for recording electromyogram (EMG) or in the periorbital region of the eye to record electrooculogram (EOG).
Recording
After recovery from surgery, animals were maintained in their home cage in a temperature-controlled environment (21±1 °C) with access to food and water ad libitum. Implanted animals demonstrated a normal behavioral repertoire immediately after recovery from surgery. However, to allow normal sleep patterns to be re-established, animals were utilized after a 3-week period. The environmental conditions described above were maintained throughout the sleep studies. For the duration of the test period, freely moving animals remained in their home cages on individual receivers. EEG and EMG or EOG signals were recorded continuously using DSI Dataquest A.R.T. The EEG trace, divided into 10-s epochs, was digitally transformed (FFT transformation) to provide the power spectra of δ, θ, α, and β bands to distinguish three different activity patterns in the rat (awake, NREM sleep, and REM sleep). The markers assigned by the automated scoring system (Sleep stage, DSI) were transferred to the EEG digital signal and subsequently confirmed by visual examination of the EEG and EMG/EOG traces by trained operators, blind to the drug treatment. Analysis of sleep parameters included: latency to NREM sleep (time interval to the first six consecutive NREM sleep epochs after injection), latency to REM sleep (time interval to the first REM sleep epoch after injection), NREM sleep, REM sleep, and total sleep time.
Drug treatment
Drug treatments were carried out according to a randomized paired crossover design where, in separate experimental sessions, each animal received vehicle or drug treatment. Rats were treated with experimental compound or its respective vehicle, in a volume of 2 ml/kg, 6 h after switch off of the light (Circadian time (CT) 18). Recordings were made for the subsequent 3-h test period. SB-649868 was dissolved in 0.5% HPMC (hydroxy-propyl-methyl-cellulose) (w/v) in distilled water and was administered by gavage at doses of 3 and 10 mg/kg. JNJ-10397049 was dissolved in mygliol 812N, and was administered intraperitoneally at doses of 5 and 25 mg/kg. GSK1059865 was dissolved in 0.5% HPMC (w/v) in distilled water and administered intraperitoneally at doses of 5 and 25 mg/kg.
Data analysis
All data are expressed as the mean±SEM. Results were analyzed using a one-way analysis of variance (ANOVA). Post-hoc comparisons were performed with Dunnett's test. Statistical significance was set at P<0.05.
Experiment 4: Binge Eating
Animals
Female Sprague–Dawley rats (Charles River) were used. Their body weight was 225–250 g at the beginning of the experiments. Rats were acclimated to individual cages under a 12-h light/dark cycle (light on at 0800 h) with ad libitum chow and water for 2 weeks before the experiments. They were kept in a room with a constant temperature (20–22 °C) and humidity (45–55%). Rats were kept in individual cages with metallic walls; the floor and the front wall were made of metallic grid. The dimensions of the cage floor were 30 cm × 30 cm; the cage was 30 cm high. A front door (30 cm × 20 cm) composed of a metallic grid was present in the anterior wall of the cage to allow access to the inside of the cage. The remaining part of the front wall was equipped with a drinking burette.
Diet
Animals were offered standard rat food pellets, 4RF18 (Mucedola; Settimo Milanese, Milano, Italy; 2.6 kcal/g). The HPF was a paste prepared by mixing Nutella (Ferrero, Alba, Torino, Italy) chocolate cream (5.33 kcal/g; 56%, 31%, and 7% from carbohydrate, fat, and protein, respectively), grounded food pellets (4RF18; Mucedola; Settimo Milanese), and water in the following weight/weight percent ratio: 52% Nutella, 33% food pellets, and 15% water. The HPF diet had a caloric content of 3.63 kcal/g. Standard pellets were offered inside a metallic grid container that was hung on the anterior wall of the cage; it was removed from the cage to measure its weight to determine food pellet intake. HPF was offered in a coffee cup; the handle of the cup was inserted into the metallic grid of the anterior wall of the cage and fixed to the wall.
The stress procedure
For 15 min, the China coffee cup containing HPF was placed inside a metallic grid container that was hung up on the anterior wall of the cage. In these conditions, the animal was able to see the cup in which it received HPF on days 5, 6, 13, and 14 of the first two cycles, was able to see the HPF itself, and also to smell its odor. In this 15-min period, the rat engaged in repeated movements of the forepaws, head, and trunk, aimed at obtaining the HPF, but it was not able to reach it.
This generated a mildly stressful condition that causes a significant increase in serum corticosterone levels (Cifani et al, 2009). After 15 min, the cup was placed inside the cage of rats of the stress groups, so that HPF became accessible to them.
Drug treatment
On day 25, compounds or respective vehicle were given before access to HPF. SB-649868 was dissolved in 0.5% HPMC (w/v) in distilled water and was administered by gavage at doses of 1 and 3 mg/kg. Topiramate was dissolved in 0.5% HPMC (w/v) in distilled water and was administered by gavage at the dose of 60 mg/kg. JNJ-10397049 was dissolved in 0.5% HPMC (w/v) in distilled water and was administered intraperitoneally at doses of 1 and 3 mg/kg. GSK1059865 was dissolved in 0.5% HPMC (w/v) in distilled water and administered by gavage at doses of 10 and 30 mg/kg. All drugs or their vehicle were administered 1 h before access to HPF.
Experiment 4A: Effect of SB-649868 and Topiramate
To evaluate the role of OXR antagonists in BE, the non-selective OXR antagonist, SB-649868, was tested in our BE model.
The animals were divided into four groups of 27 animals each, matched for body weight and daily food intake: (1) the non-restricted and not exposed to stress (NR+NS) group; (2) the restricted and not exposed to stress (R+NS) group; (3) the non-restricted and exposed to stress (NR+S) group; and (4) the restricted and exposed to stress (R+S) group. Once assigned to one of these groups, the rats remained in that group throughout the study. The rats exposed to stress were acclimated in a room different from that of the groups not exposed to stress. Rats were exposed to three consecutive 8-day cycles followed by the final test on day 25 (Table 1):
-
a)
the NR+NS group had chow ad libitum for 4 days, on days 5 and 6 they received chow+HPF for 2 h; on days 7 and 8 they had chow ad libitum; and on day 25 they were not exposed to stress;
-
b)
the second group had chow and HPF as NR+NS, but on the test day (day 25) they were exposed to stress (NR+S);
-
c)
the third group (R+NS) had chow restricted to 66% of the normal intake for 4 days, they were offered chow and HPF (2 h) on days 5 and 6 and only chow on days 7 and 8; on day 25 they were not exposed to stress.
-
d)
the R+S group had chow restricted to 66% of the normal intake for 4 days, they were offered chow and HPF (2 h) on days 5 and 6 and only chow on days 7 and 8; and on day 25 they were exposed to stress.
The 8-day cycle was repeated three times, but in the third cycle the animals did not have access to HPF food.
On day 25, each group of 27 rats was divided into three subgroups and treated, respectively, with vehicle, SB-649868, 1 or 3 mg/kg, given by gavage 1 h before access to HPF.
HPF intake was expressed as mean kcal/kg ingested±SEM; HPF intake was measured at 15, 30, 60, and 120 min after the beginning of access. Food pellet intake was measured only at 120 min, because of the findings of previous studies showing that food pellet intake was very small, and to avoid disturbance to the animals during the test.
Topiramate, used as reference compound for this experimental paradigm (Cifani et al, 2009), was tested in the same rats, 10 days after the end of SB-649868 experiment. Of these 108 animals, 72 were divided into the same four groups (18 animals for each group) described before. After one day off at the end of the first test, these groups of rats received an additional 8-day cycle: NR+NS and NR+S groups had 8 days of chow ad libitum, whereas R+NS and R+S groups had 4 days chow restricted to 66% of the normal intake followed by 4 days of chow ad libitum. In this additional cycle, all groups did not have access to HPF. The following day, NR+S and R+S groups were exposed to stress, while NR+NS and R+NS groups were not. On this day, topiramate (60 mg/kg) or its vehicle was administered by gavage 1 h before access to HPF.
Experiment 4B: Effect of JNJ-10397049 and GSK1059865
To investigate the OXR involved in reducing BE episodes, the selective OX2R antagonist, JNJ-10397049, and the selective OX1R antagonist, GSK1059865, were tested in our BE model.
An additional 54 female rats, divided into two groups (NR+NS and R+S) of 27 rats, were submitted to the same experimental procedure as in Experiment 4A. Only two groups of rats were used in this experiment as both NR+S and R+NS rats do not show BE. On the test day (day 25), 1 h before access to HPF, rats were treated intraperitoneally with JNJ-10397049 (1 and 3 mg/kg) or its vehicle.
After one day off at the end of the JNJ-10397049 test, the same group of rats received an additional 8-day cycle followed on day 10 (as described in the previous experiment) by GSK1059865 treatment. GSK1059865 (10 and 30 mg/kg) or its vehicle were administered by gavage 1 h before access to HPF.
Data analysis
All data are expressed as the mean±SEM and each value reflects the mean of number of animals per group as described in the legends. Data were analyzed by two-way ANOVA with between-subject comparisons for experimental groups or drug treatments, and within-subject comparison for time of observation. Post-hoc comparisons were carried out by the Bonferroni test. Statistical significance was set at P<0.05.
RESULTS
Experiment 1: Antagonism of SB-649868, JNJ-10397049, and GSK1059865 at rOX1R and rOX2R
OXA (0.1 nM–10 μM) increased the [3H]IPs accumulation in a concentration-dependent manner with a pEC50 value of 7.79±0.04 (n=16) and 7.68±0.04 (n=16) at rOX1R and rOX2R, respectively. At rOX1R, JNJ-10397049 (1 μM–33 μM; Figure 1a) and GSK1059865 (0.3 nM–10 nM; Figure 2a) produced non-surmountable antagonism with a dose-dependent rightward shift of the OXA EC50 and a concomitant decrease of the agonist maximal response. The calculated pKB values were 5.73±0.16 (n=3) and 8.77±0.12 (n=3) for JNJ-10397049 and GSK1059865, respectively. SB-649868 (0.1, 0.3, 0.6, and 1 nM) (Figure 3a) produced a significant reduction of OXA maximal response, without a shift of the agonist EC50. The estimated pIC50 value was 9.46±0.02 (n=3). At rOX2R JNJ-10397049 (10 nM–0.3 μM) (Figure 1b) and GSK1059865 (0.1–3.3 μM) (Figure 2b) produced a classical surmountable profile with parallel rightward shift of the OXA EC50 without depression of the agonist maximal response. The slopes obtained by Schild's regression analysis were 1.17 (95% CL 0.92–1.42) and 0.86 (95% C.L 0.71–1.00) for JNJ-10397049 and GSK1059865, respectively, and they were not statistically different from one (P>0.05). Constraining the slopes to one, the pKB values were 8.49 (95% CL 8.34–8.63; n=3) and 6.90 (95% CL 6.80–6.99; n=3) for JNJ-10397049 and GSK1059865, respectively. SB-649868 (0.1–3.3 nM) (Figure 3b) produced a dose-dependent rightward shift of the OXA EC50, accompanied by a reduction of the agonist maximal response. By applying the operational model for non-competitive antagonism analysis, as described in Materials and Methods, a pKB value of 9.35±0.15 (n=3) was calculated.

[3H]inositol phosphates (IPs) accumulation elicited by concentration–response curves (CRC) of orexin-A (OXA) (•) in rat basophil leukemia (RBL) cells expressing: rat OX1R (rOX1R) (a) in the presence of 1 (Δ), 3.3 (▾), 10 (⋄) and 33 μM (▪) of JNJ-10397049 or rOX2R (b) in the presence of 10 nM (□), 3.3 nM (Δ), 0.1 μM (▾), 0.3 μM (⋄) of JNJ-10397049. The inset graph represents the Schild's linear regression analysis of rOX2R data. Each point represents the mean±SEM of three independent experiments, each performed in duplicate.

[3H]inositol phosphates (IPs) accumulation elicited by concentration–response curves (CRC) of orexin-A (OXA) (•) in rat basophil leukemia (RBL) cells expressing: rOX1R (a) in the presence of 0.3 (▴), 1 (▾), 3.3 (♦) and 10 nM (○) of GSK1059865 or rOX2R (b) in the presence of 0.1 (□), 0.3 (Δ), 1 (▾), 3.3 μM (⋄) of GSK1059865. The inset graph represents the Schild's linear regression analysis of rOX2R data. Each point represents the mean±SEM of three independent experiments, each performed in duplicate.

[3H]inositol phosphates (IPs) accumulation elicited by concentration–response curves (CRC) of orexin-A (OXA) (•) in rat basophil leukemia (RBL) cells expressing: rat rOX1R (rOX1R) (a) in the presence of 0.1 (□), 0.3 (Δ), 0.6 (▾), and 1 nM (▪) of SB-649868 or rOX2R (b) in the presence of 0.1 (□), 0.3 (Δ), 1 (▾), and 3.3 nM (▪) of SB-649868. Each point represents the mean±SEM of three independent experiments performed in duplicate.
Experiment 2: PK Determinations in Male and Female Rats
PK determinations in male rats were investigated in the same experimental conditions as Experiment 3, and they are shown in Table 2.
Cmax of SB-649868 at 3 mg/kg was 333±52 ng/ml and AUC 1260±262 ng*h/ml.
Cmax of JNJ-10397049 at 5 mg/kg was 14.2±1.0 ng/ml and AUC 64±4.3 ng*h/ml.
Cmax of GSK1059865 at 10 mg/kg was 366±70 ng/ml and AUC 1290±320 ng*h/ml.
PK determinations in female rats were investigated in the same experimental conditions as Experiment 4, and they are shown in Table 3.
Cmax of SB-649868 at 3 mg/kg was 572±115 ng/ml and AUC 1708±331 ng*h/ml.
Cmax of JNJ-10397049 at 10 mg/kg was 369±97 ng/ml and AUC 457±224 ng*h/ml.
Cmax of GSK1059865 at 10 mg/kg was 268±29 ng/ml and AUC 768±46 ng*h/ml.
Experiment 3: Effect of SB-649868, JNJ-10397049, and GSK1059865 on Rat Sleep Model
The hypnotic profile of the OXR antagonists was evaluated over a 3-h period in the active phase of the rat, starting the recording phase at CT 18 (lights off at CT 12) of the rat light–dark cycle. CT 18 was specifically chosen to allow a maximal window to assess the hypnotic effects of compounds.
In these experimental conditions, the dual OX1/OX2R antagonist SB-649868 (3 and 10 mg/kg, by gavage) induced a robust decrease in wakefulness (F(2, 21)=22.9; P<0.01), and a decrease in sleep latency (F(2, 21)=9.11; P<0.01) compared with the control group. The Dunnett's test after ANOVA showed a statistically significant effect at 3 and 10 mg/kg (P<0.01) in both sleep parameters. The analysis of sleep patterns indicates increases in both NREM and REM sleep at both doses (Table 4a).
The selective OX2R antagonist JNJ-10397049 (5 and 25 mg/kg, intraperitoneally) showed an hypnotic effect, as revealed by the decrease of wakefulness (F(2, 14)=18.8; P<0.01) and sleep latency (F(2, 14)=4.8; P<0.05). The Dunnett's test following ANOVA showed that the decrease was statistically significant at both 5 and 25 mg/kg (P<0.01 for wakefulness and P<0.05 for sleep latency). An increase in NREM sleep was observed at both doses (P<0.01), but no effect was observed on REM sleep (Table 4b).
The selective OX1R antagonist GSK1059865 (5 and 25 mg/kg, intraperitoneally) produced a trend towards reduction of wakefulness (F(2, 14)=3.27; P<0.05) and sleep latency (F(2, 20)=1.73; P>0.05). The analysis of sleep patterns showed an increase only in NREM sleep, statistically significant at the highest tested dose (P<0.05 Dunnett's test following ANOVA); no effect was observed in REM sleep (Table 4c).
Experiment 4A. BE: Effect of SB-649868 and Topiramate
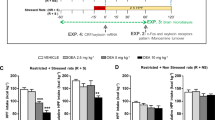
The ANOVA revealed a highly significant difference in 2-h HPF intake in the four groups of rats following vehicle administration (F(3, 32)=13.81; P<0.01). As shown in Figure 4, following vehicle administration HPF intake in the R+S group was markedly higher than that of the control (NR+NS) group. HPF intake of R+S rats was very pronounced in the first 15 min of access to it; HPF intake of the NR+S group was not significantly different from that of controls (NR+NS), indicating that stress was not sufficient to induce BE. Moreover, HPF intake of the R+NS group was not significantly different from that of controls (NR+NS), indicating that cycles of food restriction are not enough to induce BE. Therefore, BE may be caused by a unique interaction between dieting and stress.

Effect of SB-649868 (1 and 3 mg/kg, by gavage) or its vehicle on highly palatable food (HPF) intake. Values are expressed as mean±SEM of nine rats. **P<0.01, difference from vehicle-treated rats; where not indicated, the difference was not statistically significant.
The intake of standard food pellets was very small (about 3–4% of the overall calories intake in the 2 h test), and it was not affected by food restriction, stress, or by the combination of both.
As shown in Figure 4, SB-649868 significantly reduced HPF intake in the R+S group (F(2, 24)=18.63; P<0.01), but not in the other groups: NR+NS (F(2, 24)=0.91; P>0.05); R+NS (F(2, 24)=0.16; P>0.05); NR+S (F(2, 24)=1.1; P>0.05). Post-hoc comparisons revealed that the effect of SB-649868 in the R+S group was statistically significant at all the time points in response to the highest dose of 3 mg/kg. The dose of 1 mg/kg in the group R+S showed a trend towards reduction that was not statistically significant.
The topiramate test revealed a highly significant difference in 2-h HPF intake (F(3, 32)=3.93; P<0.01) of the different groups following vehicle administration. Topiramate administered at 60 mg/kg selectivity reduced HPF intake (F(1, 16)=6.57; P<0.01) in the R+S group (Figure 5), but not in NR+NS, NR+S, and R+NS groups. Data of the NR+S and R+NS groups are not shown.

Effect of topiramate (60 mg/kg, by gavage) or its vehicle on highly palatable food (HPF) intake. Values are expressed as mean±SEM of nine rats. *P<0.05, difference from vehicle treated rats; where not indicated, the difference was not statistically significant.
Experiment 4B. BE: Effect of JNJ-10397049 and GSK1059865
As for previous experiment, the ANOVA confirmed that R+S group exhibited a significant increase of HPF intake (F(1, 16)=16.17; P<0.01). JNJ-10397049 did not affect feeding either in the NR+NS group (F(2, 26)=0.23; P>0.05) or in the R+S group (F(2, 24)=0.49; P>0.05) (Figure 6a).

Effect of JNJ-10397049 (1 and 3 mg/kg, intraperitoneally) (a) or GSK1059865 (10 and 30 mg/kg, by gavage) (b) on highly palatable food (HPF) intake. Values are expressed as mean±SEM of nine rats. *P<0.05, difference from vehicle-treated rats; where not indicated, the difference was not statistically significant.
The results obtained show that OX2R antagonism did not affect feeding behavior. Hence, the effect previously observed with SB-649868 was presumably mediated through OX1R mechanisms. To confirm this finding, the selective OX1R antagonist GSK1059865 (10 and 30 mg/kg) was tested. The ANOVA showed a significant difference between the groups in response to vehicle treatment (F(1, 16)=17.1; P<0.01), confirming the BE effect in the R+S group. GSK1059865 (at the doses of 10 and 30 mg/kg) did not affect feeding in the NR+NS group (F(2, 24)=0.10; P>0.05). In the R+S group, the ANOVA showed a significant effect (F(2, 23)=4.20, P<0.05) (Figure 6b). Post-hoc comparisons revealed that the effect of GSK1059865 in the R+S group at the doses of 10 and 30 mg/kg was statistically significant at 15, 30, and 60 min after free access to HPF.
DISCUSSION
A large body of evidence suggests that dieting, stress, and negative affective states represent possible triggers of BE in patients suffering from BED or bulimia nervosa (Wardle et al, 2000; Freeman and Gil, 2004). Indeed, dieting periods are common in the histories of binge eaters, although hunger itself does not appear to be enough to induce BE in the absence of stress and negative affective state (Polivy et al, 1994; Waters et al, 2001). Considerable evidence suggests that BE may be caused by a unique interaction between dieting and stress; thus, environmental stress and a history of cyclic food restrictions may be responsible for its precipitation and maintenance (Stice et al, 2001; Crowther et al, 2001; Wolff et al, 2000). Accordingly, recurring food restrictions are consistently the strongest predictor of overeating in response to stress (Wardle et al, 2000).
The combination of dieting and stress also plays an important role in the development of BE in our preclinical model. In the model developed by Cifani et al (2009), BE is elicited by yo–yo dieting and stressful exposure to HPF. In this model, female rats are exposed to repeated cycles of restriction and a stressful procedure characterized by exposure of animals to HPF without the possibility of access to it.
As stated in the Introduction, OX mechanisms have been implicated in the control of both homeostatic and reward-based feeding, as well as in the motivation for drug of abuse (Bonci and Borgland, 2009). In keeping with the idea that neural systems that motivate and reinforce drug abuse may also underlie behaviors associated with food seeking and intake (Gearhardt et al, 2011b), this study investigated the ability of OXR antagonist to block BE episodes and to evaluate the involvement of OX1 and OX2 mechanisms in the control of BE episodes.
For this reason, three compounds reported in the literature to have different selectivity vs OX1 and OX2 human receptors were tested: the dual OX1/OX2R antagonist (SB-649868), the selective OX2R antagonist (JNJ-10397049), and the selective OX1R antagonist (GSK1059865). To use pharmacological relevant doses, the three OXR antagonists were first evaluated in vitro in rat recombinant OX1R and OX2R to determine potency and to confirm their selectivity in this animal species. The different OX1/OX2R selectivity was confirmed in rats using [3H]inositol assay.
To evaluate PK blood exposure in support of the sleep and BE studies, PK profiles of compounds have been analyzed in male and female rats, as these were the sex used in the sleep and BE experiments, respectively.
Next, the doses able to induce hypnotic effects were determined in a sleep rat model. Finally, the compounds were tested at the defined doses in the BE model.
OX1R and OX2R antagonists are reported in the literature to be involved in the control of sleep, in particular to induce hypnotic effects (Di Fabio et al, 2011; Gozzi et al, 2011; Dugovic et al, 2009). In a preclinical hypnotic sleep model in male rats, the results obtained demonstrated that the dual OX1/OX2R antagonist, SB-649868, induced a robust hypnotic effect, both on the ability to induce and to maintain sleep, reaching a statistically significant effect at 3 mg/kg. Similar to SB-649868, the OX2R antagonist JNJ-10397049 demonstrated a good hypnotic effect with a significant decrease of time spent awake at 5 mg/kg. On the contrary, the OX1R antagonist GSK1059865 showed a very poor hypnotic ability to induce and maintain sleep. These findings are in keeping with other reports suggesting that OX2R might be more important than OX1R in mediating the OX effect on sleep (Sakurai, 2007; Brisbare-Roch et al, 2007; Malherbe et al, 2009; Dugovic et al, 2009; Di Fabio et al, 2011).
SB-649868, tested in the four different conditions of stress and food restriction (NR+NS, R+NS, NR+S, R+S), was able to reduce HPF intake only in the R+S group. At 3 mg/kg, SB-649868 had an effect similar to that observed with topiramate. As with topiramate, SB-649868 did not modify HPF intake in the other conditions of food restriction and stress.
The OX2R antagonist, JNJ-10397049, tested in the same conditions did not show any effect on HPF intake in all the four tested conditions.
These results clearly indicate that the OX1R is involved in the control of BE episodes and suggested a potential role of OX1R antagonists to reverse BE episodes induced by stress and restricted diet. To verify this hypothesis, the selective OX1R antagonist GSK1059865 was evaluated in restricted and stressed animals. The results obtained confirmed that the OX1R antagonists inhibited the increase in HPF intake in R+S rats, without affecting food consumption in controls (NR+NS).
The lack of an effect of JNJ-10397049 cannot be attributed to an insufficient compound exposure in the animals submitted to the BE model. The PK evaluation of JNJ-10397049 indicated a sex difference, with females more exposed than male rats at the same dose of JNJ-10397049 (10 mg/kg). The estimated AUC values were 64 ng*h/ml in male rats vs 457 ng*h/ml in female rats (approximately seven times higher). Therefore, it is concluded that the animal exposures in the BE model at 1 and 3 mg/kg JNJ-10397049 were well above those achieved in the sleep study at 5 and 25 mg/kg. On the other hand, no sex differences were observed in the PK of both SB-649868 and GSK1059865, and exposures to the two compounds were overlapping in male and female rats.
As stated in the Introduction, stress is a key determinant of BE. Several data sets have been produced in the past decade supporting a role for OX peptides in high-arousal states, including stress, the latter being associated with substantially higher rates of OXergic neurotransmission. OX neurons in the perifornical–dorsomedial hypothalamus have been proposed to mediate stress activation (Harris and Aston-Jones, 2006, for a review, see Koob, 2008). Possibly, OXA from this hypothalamic area activates CRF-expressing neurons in the paraventricular nucleus of the hypothalamus and in the central nucleus of the amygdala (Sakamoto et al, 2004). Accordingly, OX1R antagonists inhibit reinstatement of ethanol and sucrose seeking induced by the pharmacological stressor yohimbine (Richards et al, 2008) and OXA reinstates cocaine-seeking behavior (Boutrel et al, 2005). Recently, Kuwaki (2011) found that the OX system is one of the essential modulators in the neural circuits controlling autonomic functions and emotional behaviors. Earlier findings by Johnson et al (2010) showed that the selective OX1R antagonist SB334867 attenuated the anxiety-like behavior, and blocked the increases in locomotion, heart rate, and blood pressure responses induced by the sodium lactate challenge in rat.
Several studies suggest that an altered regulation of striatal dopamine (DA) may exist in patients with bulimia nervosa and BED (Bello and Hajnal, 2010; Broft et al, 2011; Wang et al, 2011). The studies of Hoebel and co-workers (for a review, see Avena and Bocarsly, 2011) have revealed changes in striatal DA release and receptor binding, similar to those seen in response to drugs of abuse. Neuropeptides that are produced in the LH can modulate the activity of VTA–DA and striatal neurons. OX-containing neurons project from the LH to the VTA, where the OX1R plays a key role in regulating mesolimbic DA transmission and the rewarding properties of various drugs of abuse and food (Cason et al, 2010; Uramura et al, 2001; Zheng et al, 2007). Furthermore, BE episodes may be controlled through a specific influence on reward and reinforcement processes for HPF. In this regard, it is interesting to note that OX neurons in the LH have been proposed to mediate reward activation (for a review, see Koob, 2008). Thus, it is suggested that OX neurons in the LH are activated by cues associated with rewards, such as food or drugs, and stimulation of OX neurons in the LH reinstates drug seeking in rats (Harris et al, 2005).
Our group has recently demonstrated through a preclinical MRI approach that OX1R rather than OX2R selectively modulate mesolimbic brain region and the cortical part of the insula, areas implicated in rewarding processing (Gozzi et al, 2011). These data confirm and extend the previous findings that OX1R plays a role in reward processing and precipitation of drug-seeking behavior (Boutrel et al, 2005; Lawrence et al, 2006; Hollander et al, 2008; Smith et al, 2010). Thus, further work will be needed to evaluate whether the suppressive effect on BE evoked by OX1R antagonists is related to their influence on stress or reward mechanisms, or on both.
In conclusion, the results obtained in this study using three OXR antagonists with different selectivity for OX1R vs OX2R clearly demonstrated the different role of OXR in the control of BE episodes and in the modulation of sleep. Our data confirmed the principal role of OX2R mechanisms in sleep control. In addition, they show for the first time that OX1R mechanisms play a major role in the control of BE episodes. These findings suggest that targeting OX1R could represent an interesting novel pharmacological approach to the treatment of BE-related disorders.
References
American Psychiatric Association (2000). Diagnostic and Statistic Manual of Mental Disorders, IV-TR. American Psychiatric Association: Washington, DC.
Appolinario JC, Godoy-Matos A, Fontenelle LF, Carraro L, Cabral M, Vieira A et al (2000). An open-label trial of sibutramine in obese patients with binge-eating disorder. J Clin Psychiatry 63: 8–30.
Arunlakshana O, Schild HO (1959). Some quantitative uses of drug antagonists. Br J Pharmacol Chemother 14: 48–58.
Avena NM, Bocarsly ME (2011). Dysregulation of brain reward systems in eating disorders: neurochemical information from animal models of binge eating, bulimia nervosa, and anorexia nervosa. Neuropharmacology; e-pub ahead of print 27 November 2011.
Avena NM, Rada P, Hoebel BG (2008). Evidence for sugar addiction: behavioral and neurochemical effects of intermittent, excessive sugar intake. Neurosci Biobehav Rev 32: 20–39.
Bello NT, Hajnal A (2010). Dopamine and binge eating behaviors. Pharmacol Bioichem Behav 97: 25–33.
Bonci A, Borgland S (2009). Role of orexin/hypocretin and CRF in the formation of drug-dependent synaptic plasticity in the mesolimbic system. Neuropharmacology 56: 107–111.
Borgland SL, Taha SA, Sarti F, Fields HL, Bonci A (2006). Orexin A in the VTA is critical for the induction of synaptic plasticity and behavioral sensitization to cocaine. Neuron 49: 589–601.
Boutrel B, Kenny PJ, Specio SE, Martin-Fardon R, Markou A, Koob GF et al (2005). Role for hypocretin in mediating stress-induced reinstatement of cocaine-seeking behavior. Proc Natl Acad Sci USA 102: 19168–19173.
Brandish PE, Hill LA, Zheng W, Scolnick EM (2003). Scintillation proximity assay of inositol phosphates in cells extracts: high-throughput measurement of G-protein-coupled receptor activation. Anal Biochem 313: 311–318.
Brisbare-Roch C, Dingemanse J, Koberstein R, Hoever P, Aissaoui H, Flores S et al (2007). Promotion of sleep by targeting the orexin system in rats, dogs and humans. Nat Med 13: 50–155.
Broft AI, Berner LA, Martinez D, Walsh BT (2011). Bulimia nervosa and evidence for striatal dopamine dysregulation: a conceptual review. Physiol Behav 104: 122–127.
Carter WP, Hudson JI, Lalonde JK, Pindyck L, Mcelroy SL, Pope Jr HG (2003). Pharmacologic treatment of binge eating disorder. Int J Eat Disord 34 (Suppl): S74–S88.
Cason AM, Smith RJ, Tahsili-Fahadan P, Moorman DE, Sartor GC, Aston-Jones G (2010). Role of orexin/hypocretin in reward seeking and addiction: implications for obesity. Physiol Behav 100: 419–428.
Chemelli RM, Willie JT, Sinton CM, Elmquist JK, Scammell T, Lee C et al (1999). Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell 98: 437–451.
Cifani C, Polidori C, Melotto S, Ciccocioppo R, Massi MA (2009). Preclinical model of binge-eating elicited by yo–yo dieting and stressful exposure to food: effect of sibutramine, fluoxetine, topiramate and midazolam. Psychopharmacology 204: 113–125.
Corwin R, Avena NM, Boggiano MM (2011). Feeling and reward: perspective from three rat models of binge eating. Physiol Behav 104: 87–97.
Crowther JH, Sanftner J, Bonifazi DZ, Shepherd KL (2001). The role of daily hassles in binge eating. Int J Eat Disord 29: 449–454.
de Lecea L, Kilduff TS, Peyron C, Gao X, Foye PE, Danielson PE et al (1998). The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci USA 95: 322–327.
Devlin MJ, Yanovski SZ, Wilson GT (2000). Obesity: what mental health professionals need to know. Am J Psychiatry 157: 854–866.
Di Fabio R, Pellacani A, Faedo S, Roth A, Piccoli L, Gerrard P et al (2011). Discovery process and pharmacological characterization of a novel dual orexin 1 and orexin 2 receptor antagonist useful for treatment of sleep disorders. Biorg Med Chem Lett 21: 5562–5567.
Dugovic C, Shelton JE, Aluisio LE, Fraser IC, Jiang X, Sutton SW et al (2009). Blockade of orexin-1 receptors attenuates orexin-2 receptor antagonism-induced sleep promotion in the rat. J Pharmacol Exp Ther 330: 142–151.
Fassino S, Leombruni P, Pierò A, Abbate-Daga G, Giacomo Rovera G (2003). Mood, eating attitudes, and anger in obese women with and without binge eating disorder. J Psychosom Res 54: 559–566.
Foulds Mathes W, Brownley KA, Mo X, Bulik CM (2009). The biology of binge eating. Appetite 52: 545–553.
Freeman LM, Gil KM (2004). Daily stress, coping, and dietary restraint in binge eating. Int J Eat Disord 36: 204–212.
Gearhardt AN, Corbin WR, Brownell KD (2011a). Food addiction: an examination of the diagnostic criteria for dependence. J Addict Med 3: 1–7.
Gearhardt AN, Yokum S, Orr PT, Stice E, Corbin WR, Brownell KD (2011b). Neural correlates of food addiction. Arch Gen Psychiatry 68: 808–816.
Gold MS, Frost-Pineda K, Jacobs WS (2003). Overeating, binge eating, and eating disorders as addictions. Psychiatr Ann 33: 112–116.
Gozzi A, Turrini G, Piccoli L, Massagrande M, Amantini D, Antolini M et al (2011). Functional magnetic resonance imaging reveals different neural substrates for the effects of orexin-1 and orexin-2 receptor antagonists. PLoS One 6: e16406.
Grucza RA, Przybeck TR, Cloninger CR (2007). Prevalence and correlates of binge eating disorder in a community sample. Compr Psychiatry 48: 124–131.
Harris GC, Aston-Jones G (2006). Arousal and reward: a dichotomy in orexin function. Trends Neurosci 29: 571–577.
Harris GC, Wimmer M, Aston-Jones G (2005). A role for lateral hypothalamic orexin neurons in reward seeking. Nature 437: 556–559.
Haynes AC, Jackson B, Chapman H, Tadayyon M, Johns A, Porter RA et al (2000). A selective orexin-1 receptor antagonist reduces food consumption in male and female rats. Regul Pept 96: 45–51.
Heath AC (1998). Binge-eating and bulimia: potential insights into etiology and pathophysiology through genetic epidemiologic studies. Biol Psychiatry 44: 1208–1209.
Hoebel BG (1985). Brain neurotransmitters in food and drug reward. Am J Clin Nutr 42 (Suppl): 1133–1150.
Hollander JA, Lu Q, Cameron MD, Kamenecka TM, Kenny PJ (2008). Insular hypocretin transmission regulates nicotine reward. Proc Natl Acad Sci USA 105: 19480–19485.
Hudson JI, Hiripi E, Pope Jr HG, Kessler RC (2007). The prevalence and correlates of eating disorders in the national comorbidity survey replication. Biol Psychiatry 61: 348–358.
Ifland JR, Preuss HG, Marcus MT, Rourke KM, Taylor WC, Burau K et al (2009). Refined food addiction: a classic substance use disorder. Med Hypotheses 72: 518–526.
Kenakin T, Jenkinson S, Watson C (2006). Determining the potency and molecular mechanism of action of unsurmountable antagonists. J Pharmacol Exp Ther 319: 710–723.
Kenny PJ (2011). Common cellular and molecular mechanisms in obesity and drug addiction. Nat Rev Neurosci 12: 638–651.
Koob GF (2008). A role for brain stress system in addiction. Neuron 59: 11–34.
Kuwaki T (2011). Orexin links emotional stress to autonomic functions. Auton Neurosci 161: 20–27.
Javaras KN, Pope HG, Lalonde JK, Roberts JL, Nillni YI, Laird NM et al (2008). Co-occurrence of binge eating disorder with psychiatric and medical disorders. J Clin Psychiatry 269: 266–273.
Johnson PL, Truitt W, Fitz SD, Minick PE, Dietrich A, Sanghani S et al (2010). A key role for orexin in panic anxiety. Nat Med 16: 111–115.
Johnson PM, Kenny PJ (2010). Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat Neurosci 13: 635–641.
Jupp B, Krivdic B, Krstew E, Lawrence AJ (2011). The orexin1 receptor antagonist SB-334867 dissociates the motivational properties of alcohol and sucrose in rats. Brain Res 1391: 54–59.
Lawrence AJ, Cowen MS, Yang HJ, Chen F, Oldfield B (2006). The orexin system regulates alcohol-seeking in rats. Br J Pharmacol 148: 752–759.
Lu XY, Bagnol D, Burke S, Akil H, Watson SJ (2000). Differential distribution and regulation of OX1 and OX2 orexin/hypocretin receptor messenger RNA in the brain upon fasting. Horm Behav 37: 335–344.
Malherbe P, Borroni E, Pinard E, Wettstein JG, Knoflach F (2009). Biochemical and electrophysiological characterization of almorexant, a dual orexin 1 receptor (OX1)/orexin 2 receptor (OX2) antagonist: comparison with selective OX1 and OX2 antagonists. Mol Pharmacol 76: 618–631.
Marcus JN, Aschkenasi CJ, Lee CE, Chemelli RM, Saper CB, Yanagisawa M et al (2001). Differential expression of orexin receptors 1 and 2 in the rat brain. J Comp Neurol 435: 6–25.
Martin-Fardon R, Zorrilla EP, Ciccocioppo R, Weiss F (2010). Role of innate and drug-induced dysregulation of brain stress and arousal systems in addiction: focus on corticotropin-releasing factor, nociceptin/orphanin FQ, and orexin/hypocretin. Brain Res 1314: 145–161.
McAtee LC, Sutton SW, Rudolph DA, Li X, Aluisio LE, Phuong VK et al (2004). Novel substituted 4-phenyl-[1,3]dioxanes: potent and selective orexin receptor 2 (OX2R) antagonists. Bioorg Med Chem Lett 14: 4225–4229.
McElroy SL, Guerdjikova AI, Martens B, Keck Jr PE, Pope HG, Hudson JI (2009). Role of antiepileptic drugs in the management of eating disorders. CNS Drugs 23: 139–156.
McElroy SL, Hudson JI, Capece JA, Beyers K, Fisher AC, Rosenthal NR (2007). Topiramate for the treatment of binge eating disorder associated with obesity: a placebo controlled study. Biol Psychiatry 61: 1039–1048.
Nair SG, Golden SA, Shaham Y (2008). Differential effects of the hypocretin 1 receptor antagonist SB 334867 on high-fat food self-administration and reinstatement of food seeking in rats. Br J Pharmacol 154: 406–416.
Pelchat ML, Johnson A, Chan R, Valdez J, Ragland JD (2004). Images of desire: food-craving activation during fMRI. NeuroImage 23: 1486–1493.
Perello M, Sakata I, Birnbaum S, Chuang JC, Osborne-Lawrence S, Rovinsky SA et al (2010). Ghrelin increases the rewarding value of high fat diet in an orexin-dependent manner. Biol Psych 67: 880–886.
Polivy J, Zeitlin SB, Herman CP, Beal AL (1994). Food restriction and binge eating: a study of former prisoner of war. J Abnorm Psychol 103: 409–411.
Read KD, Braggio S (2010). Assessing brain free fraction in early drug discovery. Expert Opin Drug Metab Toxicol 6: 337–344.
Richards JK, Simms JA, Steensland P, Taha SA, Borgland SL, Bonci A et al (2008). Inhibition of orexin-1/hypocretin-1 receptors inhibits yohimbine-induced reinstatement of ethanol and sucrose seeking in Long–Evans rats. Psychopharmacology 199: 109–117.
Rodgers RJ, Halford JC, Nunes de Souza RL, Canto de Souza AL, Piper DC, Arch JR et al (2001). SB-334867, a selective orexin-1 receptor antagonist, enhances behavioural satiety and blocks the hyperphagic effect of orexin-A in rats. Eur J Neurosci 13: 1444–1452.
Sakamoto F, Yamada S, Ueta Y (2004). Centrally administered orexin-A activates corticotropin-releasing factor containing neurons in the hypothalamic paraventricular nucleus and central amygdaloid nucleus of rats: possible involvement of central orexins on stress-activated central CRF neurons. Reg Pept 118: 183–191.
Sakurai T (2007). The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness. Nat Rev Neurosci 8: 171–181.
Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H et al (1998). Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 92: 573–585.
Sharf R, Sarhan M, Brayton CE, Guarnieri DJ, Taylor JR, DiLeone RJ (2010). Orexin signaling via OX1R mediates operant responding for food reinforcement. Biol Psychiatry 67: 753–760.
Shoblock JR, Welty N, Aluisio L, Fraser I, Motley ST, Morton K et al (2011). Selective blockade of the orexin-2 receptor attenuates ethanol self-administration, place preference, and reinstatement. Psychopharmacology 215: 191–203.
Smith RJ, Tahsili-Fahadan P, Aston-Jones G (2010). Orexin/hypocretin is necessary for context-driven cocaine-seeking. Neuropharmacology 58: 179–184.
Stice E, Agras WS, Telch FC, Halmi KA, Mitchell JE, Wilson T (2001). Subtyping binge eating-disordered women along dieting and negative affect dimensions. Int J Eat Disord 30: 11–27.
Trivedi P, Yu H, MacNeil DJ, Van der Ploeg LH, Guan XM (1998). Distribution of orexin receptor mRNA in the rat brain. FEBS Lett 438: 71–75.
Uramura K, Funahashi H, Muroya S, Shioda S, Takigawa M, Yada T (2001). Orexin-a activates phospholipase C and protein kinase C mediated Ca2+ signaling in dopamine neurons of the ventral tegmental area. NeuroReport 12: 1885–1889.
Volkow ND, Wise RA (2005). How can drug addiction help us understand obesity? Nat Neurosci 8: 555–560.
Yager J (2008). Binge eating disorder: the search for better treatments. Am J Psychiatry 165: 4–6.
Yanovski SZ (2003). Binge eating disorder and obesity in 2003: could treating an eating disorder have a positive effect on the obesity epidemic? Int J Eat Disord 34 (Suppl): S117–S120.
Walsh BT, Devlin MJ (1998). Eating disorders: progress and problems. Science 280: 1387–1390.
Wang GJ, Geliebter A, Volkow ND, Telang FW, Logan J, Jayne MC et al (2011). Enhanced striatal dopamine release during food stimulation during binge eating disorder. Obesity 19: 1601–1608.
Wardle J, Steptoe A, Oliver G, Lipsey Z (2000). Stress, dietary restraint and food intake. J Psychosom Res 48: 195–202.
Waters A, Hill A, Waller G (2001). Internal and external antecedents of binge eating episodes in a group of women with bulimia nervosa. Int J Eat Disord 29: 17–22.
Wilfley DE, Crow SJ, Hudson JI, Mitchell JE, Berkowitz RI, Blakesley V et al (2008). Sibutramine eating disorder research group. Efficacy of sibutramine for the treatment of binge eating disorder: a randomized multicenter placebo-controlled double-blind study. Am J Psychiatry 165: 51–58.
Willie JT, Chemelli RM, Sinton CM, Yanagisawa M (2001). To eat or to sleep? Orexin in the regulation of feeding and wakefulness. Annu Rev Neurosci 24: 429–458.
Wolff GE, Crosby RD, Roberts JA, Wittrock DA (2000). Differences in daily stress, mood, coping, and eating behavior in binge eating and nonbinge eating college women. Addict Behav 25: 205–216.
Zheng H, Patterson LM, Berthoud HR (2007). Orexin signaling in the ventral tegmental area is required for high-fat appetite induced by opioid stimulation of the nucleus accumbens. J Neurosci 27: 11075–11082.
Acknowledgements
We thank Dr Charles Pickens for stylistic correction of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
EM-P is full-time employee of GSK.
Rights and permissions
About this article
Cite this article
Piccoli, L., Micioni Di Bonaventura, M., Cifani, C. et al. Role of Orexin-1 Receptor Mechanisms on Compulsive Food Consumption in a Model of Binge Eating in Female Rats. Neuropsychopharmacol 37, 1999–2011 (2012). https://doi.org/10.1038/npp.2012.48
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2012.48
Keywords
This article is cited by
-
Estradiol-dependent hypocretinergic/orexinergic behaviors throughout the estrous cycle
Psychopharmacology (2023)
-
Orexin 2 receptor (OX2R) protein distribution measured by autoradiography using radiolabeled OX2R-selective antagonist EMPA in rodent brain and peripheral tissues
Scientific Reports (2022)
-
Sleep dysregulation in binge eating disorder and “food addiction”: the orexin (hypocretin) system as a potential neurobiological link
Neuropsychopharmacology (2021)
-
Differential importance of nucleus accumbens Ox1Rs and AMPARs for female and male mouse binge alcohol drinking
Scientific Reports (2021)
-
Oleoylethanolamide decreases frustration stress-induced binge-like eating in female rats: a novel potential treatment for binge eating disorder
Neuropsychopharmacology (2020)
