
Abstract
Some plants acquire competence to flower in spring after experiencing a seasonal temperature drop—winter cold, in a process termed vernalization. In Arabidopsis thaliana, prolonged exposure to cold induces epigenetic silencing of the potent floral repressor locus FLOWERING LOCUS C (FLC) by Polycomb group (PcG) proteins, and this silencing is stably maintained in subsequent growth and development upon return to warm temperatures. Here we show that a cis-regulatory DNA element in the nucleation region for PcG silencing at FLC and two homologous trans-acting epigenome readers, VAL1 and VAL2, control vernalization-mediated FLC silencing. The sequence-specific readers recognize both the cis element (termed the cold memory element) and a repressive mark, trimethylation of histone H3 at lysine 27 (H3K27me3), and directly associate with LIKE HETEROCHROMATIN PROTEIN 1 (LHP1), leading to establishment of the H3K27me3 peak in the nucleation region at FLC during vernalization. Thus, our work describes a mechanism for PcG-mediated silencing by a DNA sequence-specific epigenome reader.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

References
Lanzuolo, C. & Orlando, V. Memories from the Polycomb group proteins. Annu. Rev. Genet. 46, 561–589 (2012).
Meller, V.H., Joshi, S.S. & Deshpande, N. Modulation of chromatin by noncoding RNA. Annu. Rev. Genet. 49, 673–695 (2015).
Berry, S. & Dean, C. Environmental perception and epigenetic memory: mechanistic insight through FLC. Plant J. 83, 133–148 (2015).
Amasino, R. Seasonal and developmental timing of flowering. Plant J. 61, 1001–1013 (2010).
Helliwell, C.A., Anderssen, R.S., Robertson, M. & Finnegan, E.J. How is FLC repression initiated by cold? Trends Plant Sci. 20, 76–82 (2015).
Heo, J.B. & Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 331, 76–79 (2011).
Sung, S. & Amasino, R.M. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature 427, 159–164 (2004).
De Lucia, F., Crevillen, P., Jones, A.M., Greb, T. & Dean, C. A PHD–Polycomb repressive complex 2 triggers the epigenetic silencing of FLC during vernalization. Proc. Natl. Acad. Sci. USA 105, 16831–16836 (2008).
Angel, A., Song, J., Dean, C. & Howard, M. A Polycomb-based switch underlying quantitative epigenetic memory. Nature 476, 105–108 (2011).
Finnegan, E.J. & Dennis, E.S. Vernalization-induced trimethylation of histone H3 lysine 27 at FLC is not maintained in mitotically quiescent cells. Curr. Biol. 17, 1978–1983 (2007).
Swiezewski, S., Liu, F., Magusin, A. & Dean, C. Cold-induced silencing by long antisense transcripts of an Arabidopsis Polycomb target. Nature 462, 799–802 (2009).
Helliwell, C.A., Robertson, M., Finnegan, E.J., Buzas, D.M. & Dennis, E.S. Vernalization-repression of Arabidopsis FLC requires promoter sequences but not antisense transcripts. PLoS One 6, e21513 (2011).
Berry, S., Hartley, M., Olsson, T.S., Dean, C. & Howard, M. Local chromatin environment of a Polycomb target gene instructs its own epigenetic inheritance. eLife 4, 07205 (2015).
He, Y., Michaels, S.D. & Amasino, R.M. Regulation of flowering time by histone acetylation in Arabidopsis. Science 302, 1751–1754 (2003).
Michaels, S.D. & Amasino, R.M. FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell 11, 949–956 (1999).
Lee, I., Michaels, S.D., Masshardt, A.S. & Amasino, R.M. The late-flowering phenotype of FRIGIDA and luminidependens is suppressed in the Landsberg erecta strain of Arabidopsis. Plant J. 6, 903–909 (1994).
Suzuki, M., Kao, C.Y. & McCarty, D.R. The conserved B3 domain of VIVIPAROUS1 has a cooperative DNA binding activity. Plant Cell 9, 799–807 (1997).
Suzuki, M., Wang, H.H. & McCarty, D.R. Repression of the LEAFY COTYLEDON 1/B3 regulatory network in plant embryo development by VP1/ABSCISIC ACID INSENSITIVE 3-LIKE B3 genes. Plant Physiol. 143, 902–911 (2007).
Tsukagoshi, H., Saijo, T., Shibata, D., Morikami, A. & Nakamura, K. Analysis of a sugar response mutant of Arabidopsis identified a novel B3 domain protein that functions as an active transcriptional repressor. Plant Physiol. 138, 675–685 (2005).
King, G.J. et al. The Arabidopsis B3 domain protein VERNALIZATION1 (VRN1) is involved in processes essential for development, with structural and mutational studies revealing its DNA-binding surface. J. Biol. Chem. 288, 3198–3207 (2013).
Yamasaki, K. et al. Solution structure of the B3 DNA binding domain of the Arabidopsis cold-responsive transcription factor RAV1. Plant Cell 16, 3448–3459 (2004).
Yang, C. et al. VAL- and AtBMI1-mediated H2Aub initiate the switch from embryonic to postgerminative growth in Arabidopsis. Curr. Biol. 23, 1324–1329 (2013).
Kim, H.J. et al. A genetic link between cold responses and flowering time through FVE in Arabidopsis thaliana. Nat. Genet. 36, 167–171 (2004).
Ausín, I., Alonso-Blanco, C., Jarillo, J.A., Ruiz-García, L. & Martínez-Zapater, J.M. Regulation of flowering time by FVE, a retinoblastoma-associated protein. Nat. Genet. 36, 162–166 (2004).
Veerappan, V. et al. A novel HSI2 mutation in Arabidopsis affects the PHD-like domain and leads to derepression of seed-specific gene expression. Planta 236, 1–17 (2012).
Hoppmann, V. et al. The CW domain, a new histone recognition module in chromatin proteins. EMBO J. 30, 1939–1952 (2011).
Veerappan, V., Chen, N., Reichert, A.I. & Allen, R.D. HSI2/VAL1 PHD-like domain promotes H3K27 trimethylation to repress the expression of seed maturation genes and complex transgenes in Arabidopsis seedlings. BMC Plant Biol. 14, 293 (2014).
Sanchez, R. & Zhou, M.M. The PHD finger: a versatile epigenome reader. Trends Biochem. Sci. 36, 364–372 (2011).
Zhang, X. et al. The Arabidopsis LHP1 protein colocalizes with histone H3 Lys27 trimethylation. Nat. Struct. Mol. Biol. 14, 869–871 (2007).
Turck, F. et al. Arabidopsis TFL2/LHP1 specifically associates with genes marked by trimethylation of histone H3 lysine 27. PLoS Genet. 3, e86 (2007).
Sung, S. et al. Epigenetic maintenance of the vernalized state in Arabidopsis thaliana requires LIKE HETEROCHROMATIN PROTEIN 1. Nat. Genet. 38, 706–710 (2006).
Derkacheva, M. et al. Arabidopsis MSI1 connects LHP1 to PRC2 complexes. EMBO J. 32, 2073–2085 (2013).
Wang, Y., Gu, X., Yuan, W., Schmitz, R.J. & He, Y. Photoperiodic control of the floral transition through a distinct Polycomb repressive complex. Dev. Cell 28, 727–736 (2014).
Qüesta, J.I., Song, J., Geraldo, N., An, H. & Dean, C. Arabidopsis transcriptional repressor VAL1 triggers Polycomb silencing at FLC during vernalization. Science 353, 485–488 (2016).
Michaels, S.D. & Amasino, R.M. Loss of FLOWERING LOCUS C activity eliminates the late-flowering phenotype of FRIGIDA and autonomous pathway mutations but not responsiveness to vernalization. Plant Cell 13, 935–941 (2001).
Hajdukiewicz, P., Svab, Z. & Maliga, P. The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 25, 989–994 (1994).
Karimi, M., De Meyer, B. & Hilson, P. Modular cloning in plant cells. Trends Plant Sci. 10, 103–105 (2005).
Gu, X. et al. Arabidopsis FLC clade members form flowering-repressor complexes coordinating responses to endogenous and environmental cues. Nat. Commun. 4, 1947 (2013).
Jiang, D., Kong, N.C., Gu, X., Li, Z. & He, Y. Arabidopsis COMPASS-like complexes mediate histone H3 lysine-4 trimethylation to control floral transition and plant development. PLoS Genet. 7, e1001330 (2011).
Wysocka, J. et al. WDR5 associates with histone H3 methylated at K4 and is essential for H3 K4 methylation and vertebrate development. Cell 121, 859–872 (2005).
Acknowledgements
We thank M. Suzuki (University of Florida) for kindly providing val1 and val2 seeds and the Flanders Interuniversity Institute for Biotechnology (Belgium) for providing pBGW vector. This work was supported in part by funding from the Chinese Academy of Sciences and from the Ministry of Science and Technology of China (grant 2016YFA0503200 to J.D.).
Author information
Authors and Affiliations
Contributions
Y.H. conceived the project. W. Yuan, X.L., Z.L., W. Yang, Y.W., and R.L. performed the experiments. W. Yuan, X.L., Z.L., W. Yang, Y.W., R.L., J.D., and Y.H. analyzed the data. Y.H. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 A cis-regulatory region in the first intron of FLC mediates transcriptional repression.
(a) Relative FLC transcript levels in the indicated transgenic seedlings (in the FRI flc-3 background). A total of eight independent T1 lines for each transgene were examined. (b) The 10-bp TTCTGCATGG sequence mediates FLC repression independently of the autonomous pathway gene FCA. FCA represses FLC expression to promote flowering3. WT-FLC and FLC ∆10 (transgenes) were introduced into fca flc-3; the 10-bp deletion caused further FLC upregulation and consequent late flowering. “Non-flower” denotes plants that had not flowered after 3 months of growth in long days. (c) Relative mRNA levels of the GUS fusion with part of FLM and the 5′ UTR of SPY in the indicated T1 transgenic plants. 35–38 independent T1 seedlings from each line were pooled for RNA extraction. Error bars, s.d. of three biological repeats. (d) FLC mRNA levels in T1 plants expressing the indicated transgenes. 30, 28, and 30 independent T1 seedlings were pooled for RNA extraction from WT-FLC, FLCm1, and FLCm2, respectively. In a, c, and d, FLC levels were normalized to those of the endogenous control, TUB2. In a and d, error bars, s.d. of triplicate quantifications. (e) Box plots of the flowering times of the indicated lines (T1 generation; in the flc-2 background). Center lines represent medians, box limits represent the 25th and 75th percentiles, and whiskers represent the ranges of data; red dots are data points.
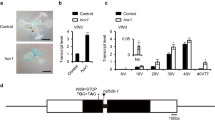
Supplementary Figure 2 Schematic of the FLC locus.
On top is the sequence of the 47-bp FLC silencing element (Sph/RY motifs are in red; numbers correspond to nucleotide positions, with A of the ATG codon as +1). Arrows indicate a transcription start site (TSS); boxes correspond to exons. VRE denotes a 289-bp region around the COLDAIR TSS, the deletion of which disrupts the maintenance of vernalization-mediated FLC repression upon return to warm temperature31. COOLAIR RNAs mainly consist of three isoforms denoted by purple31.
Supplementary Figure 3 VAL1 and VAL2, but not VAL3, RAV1, or VRN1, recognize and bind to the Sph/RY-bearing FLC silencing element.
WT-72 bp, m1.m1, and m2.m2 denote the 72-bp FLC fragment (+451 to +522, where A of the ATG codon is +1), a mutant fragment with both Sph/RY motifs mutated with the m1 mutation, and a fragment with both motifs mutated to the m2 mutation, respectively.
Supplementary Figure 4 Analyses of val1, val2, val3, and val1 val2 mutants.
(a) Box plots of the flowering times of the indicated lines in long days. Box plots show the median line, interquartile range (box limits), whiskers (extending 1.5 times the interquartile range), and data points (red cycles). Fifteen plants were scored for each genotype. (b) Phenotypes of 4-d-old val1 val2 seedlings. The selfed progeny of a weak val1 val2 double mutant exhibit a 1:9 ratio of strong to weak mutants (a total of 267 mutants were scored at an early seedling stage). Scale bars, 1 mm. (c) pVAL1-VAL1:Flag rescued the val1 val2 double-mutant phenotype. Shown are 3-week-old plants grown in long days. Scale bars, 1 cm.
Supplementary Figure 5 Analyses of COOLAIR and COLDAIR RNA levels during the course of vernalization in wild-type and val1 val2 seedlings.
(a,b) Relative COOLAIR (a) and COLDAIR (b) RNA levels. RNA levels were normalized to those of PP2A; error bars, s.d. of two biological repeats.
Supplementary Figure 6 Analysis of FLCm1 copy number by qPCR.
An FLC intron I region from wild type (Col) or the indicated transgenic lines (T3 homozygotes and in the flc-2 background) was quantified, and signal was first normalized to that of the single-copy reference gene FLOWERING LOCUS T, followed by normalization to that of the endogenous FLC in wild type. Error bars, s.d. of triplicate quantification. Both FLCm1-1 and FLCm1-2 bear a single T-DNA insertion.
Supplementary Figure 7 VAL1 and VAL2 can form homodimers.
Yeast two-hybrid assays with full-length VAL1 and VAL2 were conducted.
Supplementary Figure 8 Analysis of His-SUMO-VAL1N.
(a) Schematic of the VAL1 protein. The gray bar denotes VAL1N. (b) Affinity-purified His-SUMO-VAL1N from E. coli. Purifications were analyzed by SDS–PAGE. (c) His-SUMO-VAL1N, but not His-SUMO, binds to H3K27me2 and H3K27me3, as demonstrated by histone 3 peptide pulldown assays.
Supplementary Figure 9 Working model for the regulatory CME–VAL1/VAL2–H3K27me3 interactions to mediate Polycomb silencing of FLC by vernalization.
Before vernalization, FLC expression in early seedlings is subject to dynamic control by the active histone 3 lysine 36 methyltransferase EFS and a repressive core PRC2 complex recruited by CME–VAL1/VAL2–LHP1. During long exposure to cold, more VAL proteins (and LHP1) bind to the CME-containing nucleation region, leading to the recruitment of PHD (VIN3)–PRC2 to establish the H3K27me3 peak at this region (in one or both FLC alleles) and generate a positive feedback loop, resulting in silencing of an FLC allele. Upon return to warm temperature, the silenced FLC allele bearing repressive H3K27me3 is read by VAL1/VAL2–LHP1 through binding to both the cis element CME and H3K27me3, enabling the instruction of an allele-specific repressive chromatin state for its own inheritance during mitotic cell divisions.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–9 and Supplementary Table 1. (PDF 1045 kb)
Supplementary Data
Uncropped immunoblots. (PDF 102 kb)
Rights and permissions
About this article

Cite this article
Yuan, W., Luo, X., Li, Z. et al. A cis cold memory element and a trans epigenome reader mediate Polycomb silencing of FLC by vernalization in Arabidopsis. Nat Genet 48, 1527–1534 (2016). https://doi.org/10.1038/ng.3712
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ng.3712
This article is cited by
-
A VEL3 histone deacetylase complex establishes a maternal epigenetic state controlling progeny seed dormancy
Nature Communications (2023)
-
LHP1-mediated epigenetic buffering of subgenome diversity and defense responses confers genome plasticity and adaptability in allopolyploid wheat
Nature Communications (2023)
-
Polycomb Repressive Complexes and Their Roles in Plant Developmental Programs, Particularly Floral Transition
Journal of Plant Biology (2023)
-
HEAT SHOCK TRANSCRIPTION FACTOR B2b acts as a transcriptional repressor of VIN3, a gene induced by long-term cold for flowering
Scientific Reports (2022)
-
Vernalization Regulates Flowering Genes and Modulates Glucosinolates Biosynthesis in Chinese Cabbage
Journal of Plant Biology (2022)
