
Abstract
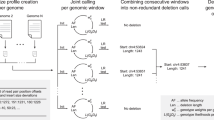
The locations and properties of common deletion variants in the human genome are largely unknown. We describe a systematic method for using dense SNP genotype data to discover deletions and its application to data from the International HapMap Consortium to characterize and catalogue segregating deletion variants across the human genome. We identified 541 deletion variants (94% novel) ranging from 1 kb to 745 kb in size; 278 of these variants were observed in multiple, unrelated individuals, 120 in the homozygous state. The coding exons of ten expressed genes were found to be commonly deleted, including multiple genes with roles in sex steroid metabolism, olfaction and drug response. These common deletion polymorphisms typically represent ancestral mutations that are in linkage disequilibrium with nearby SNPs, meaning that their association to disease can often be evaluated in the course of SNP-based whole-genome association studies.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

References
Sebat, J. et al. Large-scale copy number polymorphism in the human genome. Science 305, 525–528 (2004).
Iafrate, A.J. et al. Detection of large-scale variation in the human genome. Nat. Genet. 36, 949–951 (2004).
Sharp, A.J. et al. Segmental duplications and copy-number variation in the human genome. Am. J. Hum. Genet. 77, 78–88 (2005).
Tuzun, E. et al. Fine-scale structural variation of the human genome. Nat. Genet. 37, 727–732 (2005).
International HapMap Consortium. A haplotype map of the human genome. Nature 437, 1299–1320 (2005).
Daiger, S.P. & Chakravarti, A. Deletion mapping of polymorphic loci by apparent parental exclusion. Am. J. Med. Genet. 14, 43–48 (1983).
Chance, P.F. et al. DNA deletion associated with hereditary neuropathy with liability to pressure palsies. Cell 72, 143–151 (1993).
Bailey, J.A. et al. Recent segmental duplications in the human genome. Science 297, 1003–1007 (2002).
Seidegard, J., Vorachek, W.R., Pero, R.W. & Pearson, W.R. Hereditary differences in the expression of the human glutathione transferase active on trans-stilbene oxide are due to a gene deletion. Proc. Natl. Acad. Sci. USA 85, 7293–7297 (1988).
Nunoya, K. et al. A new deleted allele in the human cytochrome P450 2A6 (CYP2A6) gene found in individuals showing poor metabolic capacity to coumarin and (+)-cis-3,5-dimethyl-2-(3-pyridyl)thiazolidin-4-one hydrochloride (SM-12502). Pharmacogenetics 8, 239–249 (1998).
Pemble, S. et al. Human glutathione S-transferase theta (GSTT1): cDNA cloning and the characterization of a genetic polymorphism. Biochem. J. 300, 271–276 (1994).
Wilson, W. et al. Characterization of a common deletion polymorphism of the UGT2B17 gene linked to UGT2B15. Genomics 84, 707–714 (2005).
Monks, S.A. et al. Genetic inheritance of gene expression in human cell lines. Am. J. Hum. Genet. 75, 1094–1105 (2004).
Morley, M. et al. Genetic analysis of genome-wide variation in human gene expression. Nature 430, 743–747 (2004).
de Bakker, P.W. et al. Efficiency and power in genetic association studies. Nat. Genet. (in the press).
Conrad, D.F., Andrews, T.D., Carter, N.P., Hurles, M.E. & Pritchard, J.K. A high-resolution survey of deletion polymorphism in the human genome. Nat. Genet., advance online publication 4 December 2005 (10.1038/ng1697).
Hinds, D.A., Kloek, A.P. & Frazer, K.A. Common deletions and SNPs are in linkage disequilibrium in the human genome. Nat. Genet., advance online publication 4 December 2005 (10.1038/ng1695).
Barrett, J.C. et al. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 21, 263–265 (2005).
Fry, B. Computational Information Design. Thesis, Massachusetts Institute of Technology (2005).
Acknowledgements
The authors wish to thank J. Moore and L. Ziaugra for contributing their expertise on the behavior of genotyping platforms and C. Patil, J. Melo and E. Lander for commenting on manuscript drafts. We thank G. Thorisson and A. Vernon-Smith for extensive help with data coordination, and D. Conrad, J. Pritchard and K. Frazer for exchanging manuscripts before publication.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Fig. 1
Physical clustering of patterns of apparent mendelian inconsistency and null genotypes in the HapMap data. (PDF 119 kb)
Supplementary Fig. 2
Distinct patterns of aberrant SNP genotypes caused by the same deletion polymorphisms in multiple populations. (PDF 185 kb)
Supplementary Fig. 3
Confirmation of segregating deletion variants by fluorescent in situ hybridization (FISH). (PDF 1530 kb)
Supplementary Fig. 4
Deletion variants flanked by segmental duplications. (PDF 496 kb)
Supplementary Fig. 5
Linkage disequilibrium between gene deletion polymorphisms and nearby SNPs. (PDF 317 kb)
Supplementary Table 1
Predicted deletion variants and supporting SNP evidence. (PDF 230 kb)
Supplementary Table 2
Validation of candidate deletion variants. (PDF 17 kb)
Supplementary Table 3
Gene deletion genotypes obtained by quantitative PCR for ten loci in 269 individuals. (PDF 66 kb)
Supplementary Table 4
SNP alleles that tag common gene deletion alleles, for potential use in medical genetic studies. (PDF 12 kb)
Supplementary Table 5
Primer and probe sequences used. (PDF 29 kb)
Rights and permissions
About this article
Cite this article
McCarroll, S., Hadnott, T., Perry, G. et al. Common deletion polymorphisms in the human genome. Nat Genet 38, 86–92 (2006). https://doi.org/10.1038/ng1696
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ng1696
This article is cited by
-
Ethnic and functional differentiation of copy number polymorphisms in Tunisian and HapMap population unveils insights on genome organizational plasticity
Scientific Reports (2024)
-
Genetic variation in the immunoglobulin heavy chain locus shapes the human antibody repertoire
Nature Communications (2023)
-
Extensive metabolic consequences of human glycosyltransferase gene knockouts in prostate cancer
British Journal of Cancer (2023)
-
Assessment of linkage disequilibrium patterns between structural variants and single nucleotide polymorphisms in three commercial chicken populations
BMC Genomics (2022)
-
Understanding Mendelian errors in SNP arrays data using a Gochu Asturcelta pig pedigree: genomic alterations, family size and calling errors
Scientific Reports (2022)
