
Abstract
DNA methylation is an epigenetic repressor mark for transcription dynamically regulated in neurons. We analyzed visual experience regulation of DNA methylation in mice and its involvement in ocular dominance plasticity of the developing visual cortex. Monocular deprivation modulated the expression of factors controlling DNA methylation and exerted opposite effects on DNA methylation and hydroxymethylation in specific plasticity genes. Inhibition of DNA methyltrasferase (DNMT) blocked molecular and functional effects of monocular deprivation, partially reversing the monocular deprivation transcriptional program.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others

References
Moore, L.D., Le, T. & Fan, G. Neuropsychopharmacology 38, 23–38 (2013).
Tognini, P., Putignano, E., Coatti, A. & Pizzorusso, T. Nat. Neurosci. 14, 1237–1239 (2011).
Mellios, N. et al. Nat. Neurosci. 14, 1240–1242 (2011).
Hong, E.J., McCord, A.E. & Greenberg, M.E. Neuron 60, 610–624 (2008).
Remenyi, J. et al. Biochem. J. 428, 281–291 (2010).
Martinowich, K. et al. Science 302, 890–893 (2003).
Brueckner, B. et al. Cancer Res. 65, 6305–6311 (2005).
Day, J.J. et al. Nat. Neurosci. 16, 1445–1452 (2013).
LaPlant, Q. et al. Nat. Neurosci. 13, 1137–1143 (2010).
Guo, J.U. et al. Nat. Neurosci. 14, 1345–1351 (2011).
Huang, Y., Pastor, W.A., Zepeda-Martinez, J.A. & Rao, A. Nat. Protoc. 7, 1897–1908 (2012).
Pastor, W.A., Huang, Y., Henderson, H.R., Agarwal, S. & Rao, A. Nat. Protoc. 7, 1909–1917 (2012).
Szulwach, K.E. et al. Nat. Neurosci. 14, 1607–1616 (2011).
Shi, F.T. et al. J. Biol. Chem. 288, 20776–20784 (2013).
Wang, Y. & Zhang, Y. Cell Reports 6, 278–284 (2014).
Bauer, C. et al. J. Biol. Chem. 290, 4801–4812 (2015).
Xu, W. et al. Cancer Cell 19, 17–30 (2011).
Blaschke, K. et al. Nature 500, 222–226 (2013).
Yin, R. et al. J. Am. Chem. Soc. 135, 10396–10403 (2013).
Krol, J. et al. Cell 141, 618–631 (2010).
Weber, M. et al. Nat. Genet. 37, 853–862 (2005).
Kember, R.L. et al. Brain Behav. 2, 455–467 (2012).
Carmona-Saez, P., Chagoyen, M., Tirado, F., Carazo, J.M. & Pascual-Montano, A. Genome Biol. 8, R3 (2007).
Nogales-Cadenas, R. et al. Nucleic Acids Res. 37, W317–W322 (2009).
Tabas-Madrid, D., Nogales-Cadenas, R. & Pascual-Montano, A. Nucleic Acids Res. 40, W478–W483 (2012).
Lodovichi, C., Berardi, N., Pizzorusso, T. & Maffei, L. J. Neurosci. 20, 2155–2165 (2000).
Rittenhouse, C.D., Shouval, H.Z., Paradiso, M.A. & Bear, M.F. Nature 397, 347–350 (1999).
Rossi, F.M. et al. Proc. Natl. Acad. Sci. USA 98, 6453–6458 (2001).
Schuett, S., Bonhoeffer, T. & Hubener, M. J. Neurosci. 22, 6549–6559 (2002).
Acknowledgements
We thank K. Eckel-Mahan for critical reading of the manuscript, and N. Huang and A. Rao (La Jolla Institute for Allergy and Immunology) for the CMS antibody. This work was supported by Epigenomics Flagship Project EPIGEN MIUR-CNR to M.D'E. and T.P.
Author information
Authors and Affiliations
Contributions
P.T., D.N. and J.T. performed the molecular experiments. P. T. and D.S. performed the electrophysiology. D.N., M.D'E. and F.D.R. performed and supervised the bisulfite analysis. P.T. and T.P. wrote the paper. T.P. supervised the project.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Monocular deprivation regulates DNMT expression in the visual cortex of critical period mice.
(a) MD3d significantly increased the expression of Dnmt3a, Dnmt3b and Dnmt1 mRNA in the visual cortex contralateral (contra ctx) to the deprived eye with respect to the ipsilateral cortex (ipsi ctx) (cortex ipsilateral to the deprived eye vs. contralateral cortex, signed rank test, Dnmt3a n = 18, W = 5, P = 0.02; Dnmt3b n = 13, W = 79, P = 0.003; Dnmt1 n = 13, W = 89, p < 0.001). (b) Dnmt3a, Dnmt3b and Dnmt1 expression in the right (right ctx) and left cortex (left ctx) of aged matched non deprived (Nondep) animals. No significant differences were found (right ctx vs. left ctx, n = 10, signed rank test, Dnmt3a W = 5 P = 0.82; Dnmt3b W = 23, P = 0.92; Dnmt1 W = 34,P =0.21). (c) qPCR quantitation of Dnmt3a (a), Dnmt3b (b) and Dnmt1 (c) in the visual cortex of MD3d and age matched non deprived mice are expressed as percentage of Gapdh. (MD3d: ipsi ctx vs. contra ctx, Dnmt3a n = 18, W = 97; Dnmt3b n=13,W=-79;Dnmt1n=13,W=89,Signed rank test p<0.05. Nondep: n=10, Dnmt3aW= 5, P = 0.81; Dnmt3b W = -3, P = 0.92; Dnmt1 W = 34, P = 0.2) (d) Western blot analysis of DNMT3a and DNMT1 in the visual cortex of MD3d mice (DNMT3A n = 13, W = 72; DNMT1 n = 16, W = 120, signed rank test P < 0.001). Error bars represent SEM.
Supplementary Figure 2 Widespread expression of DNMT3a and DNMT1 in P27 visual cortex.
Immunohistochemistry of DNMT3a and DNMT1 in the visual cortex of critical period mice: double staining with NeuN (specific neuronal marker), GAD67 (specific inhibitory neuron marker) and GFAP (specific astroglial marker) demonstrated protein expression in all the investigated cell types.
Supplementary Figure 3 Monocular deprivation regulates GADD45 family members expression in the visual cortex of critical period mice.
MD3d significantly decreased Gadd45 family members gene expression in the cortex contralateral to the deprived eye (contra ctx) with respect to the ipsilateral cortex (ipsi ctx) (cortex ipsilateral to the deprived eye vs. contralateral cortex, Gadd45a n = 8, W = -36, signed rank test P = 0.008; Gadd45b n = 16, W = -36, P = 0.008; Gadd45g n = 16, W = -28, P = 0.048). Error bars represent SEM.
Supplementary Figure 4 Specificity of the MeDIP experiment.
(a) Sketch of miR212/132 and Bdnfex-4 CRE sites and interrogated sequences. (b) Normal rabbit IgG caused a statistically significant less recovery of methylated DNA with respect to a specific antibody against 5-mC. BDNFex-4b, BDNFex-4c and CRE2-3 sequence abundance was measured using qPCR in cortical samples immunoprecipitated with antibodies specific for 5-mC or Normal rabbit IgG. Data were normalized to the average recovery obtained with the antibody for 5-mC. (BDNFex-4b: n = 11, T = 72; BDNFex-4c T = 92, n =12; CRE2-3 n = 11, T = 75, 5-mC vs Normal rabbit IgG, Mann-Withney rank sum test p<0.001. (c) 100% methylated DNA and DNA from visual cortical samples were immunoprecipitated with purified antibody against 5-mC. 100% methylated DNA shows a significantly higher recovery in respect to the other sample. (n = 5, T = 16, Mann-Withney rank sum test P = 0.005). Error bars represent SEM.
Supplementary Figure 5 hMeDIP data obtained with a 5-hmC antiserum.
Using a 5-hmc antiserum we confirmed the results obtained with a purified antibody. MD3d induced a significant decrease in DNA hydroxylmethylation on CRE1, CRE2-3, BDNFex-4c, but not on CRE4, in the visual cortex contralateral to the deprived eye with respect to the ipsilateral cortex. 5-hmC showed a trend for decrease at the BDNFex-4b promoter. (contra ctx vs ipsi ctx, signed rank test, CRE1 n = 12, W = -50, P = 0.05; CRE2-3 n = 8, W = -22, P = 0.04; CRE4 n = 13, W = -31 P = 0.31; BDNFex4-b n = 12 W = -36, P = 0.15, BDNFex-4c n = 13, W = -53 P = 0.048). Error bars represent SEM.
Supplementary Figure 6 Specificity of the hMeDIP experiment.
Normal rabbit IgG caused a statistically significant reduced recovery of hydroxylmethylated DNA with respect to a specific antibody against 5-hmC. BDNFex-4b (a), BDNFex-4c (b) and CRE2-3 (c) sequence abundance was measured using qPCR in cortical samples immunoprecipitated with antibodies specific for 5-hmC or Normal rabbit IgG. Data were normalized to the average recovery obtained with the antibody for 5-hmC. a) BDNFex-4b: n = 11, T = 7, b) BDNFex-4c n =12, T = 78, c) CRE2-3 n = 11, T = 66, 5-hmC vs Normal rabbit IgG, Mann-Withney rank sum test p < 0.001. Error bars represent SEM.
Supplementary Figure 7 MeDIP and hMeDIP in MD3d adult mice.
MD3d in adult (p120) animals is not able to regulate DNA methylation (a) and hydroxylmethylation (b). MeDIP n = 6-7: ipsi ctx vs contra ctx, paired t- test, CRE1: t = 0.91, P = 0.4; CRE2-3: t = 0.51, P = 0.63; CRE4: t = 0.57, P = 0.53; BDNFex4-b: t = 0.73, P = 0.5; BDNFex4-c: t = 0.38, P = 0.72. hMeDIP: n = 6-8 ipsi ctx vs contra ctx, paired t-test, CRE1: t = 0.97, P = 0.37; CRE2-3: t = 0.02, P = 0.99; CRE4: t = 0.81, P = 0.44; BDNFex4-b: t = 0.92, P = 0.4; BDNFex4-c: t = 0.01, P = 0.99. Error bars represent SEM.
Supplementary Figure 8 CMS IP in MD3d critical period mice.
a) CMS technique flow chart. b) Using an anti-CMS antibody to perform the immunoprecipitation of bisulfite-converted DNA we confirmed the results obtained with hMeDIP. MD3d induced a significant decrease in DNA hydroxylmethylation on CRE1, CRE2-3, BDNFex-4b/c, in the visual cortex contralateral to the deprived eye with respect to the ipsilateral cortex (contra ctx vs ipsi ctx, paired t-test, CRE1 n = 5, t = 3.49, P = 0.025; CRE2-3 n = 5, t = 3.76, P = 0.02; BDNFex4-b/c n = 5 t = 3.43, P = 0.03). Error bars represent SEM. CMS-IP has been shown to perform equal or better than glucosylation, periodate oxidation and biotinylation (GLIB) technique or hMeDIP (Pastor, W. A., Huang, Y., Henderson, H. R., Agarwal, S. & Rao, A. The GLIB technique for genome-wide mapping of 5-hydroxymethylcytosine. Nat Protoc 7, 1909-1917, (2012); Huang, Y., Pastor, W. A., Zepeda-MarTnez, J.A. & Rao, A. The anti-CMS technique for genome-wide mapping of 5-hydroxymethylcytosine. Nat Protoc 7, 1897-1908, (2012). The anti-CMS antiserum was generously provided by Dr. Nancy Huang and Dr. Anjana Rao. Since CMS-IP requires bisulfite conversion, to assess the abundance of the DNA loci of interest in the immunoprecipitate we designed new primers insensitive to the methylation status of the complementary DNA sequences. BDNFex-4b and BDNFex-4c could be evaluated with a single primer set designated BDNFex-4b/c.
Supplementary Figure 9 Tet expression in the visual cortex of juvenile mice
(a) Tet1, Tet2 and Tet3 mRNA expression was quantified by qPCR. The binocular area in the visual cortex of MD3d critical period mice and age matched control Nondep animals was analyzed. Tet1 MD3d n = 17, ipsi ctx vs contra ctx, paired t-test t = -0.278, P = 0.78; Nondep n = 10, t = 1.06, P = 0.32. Tet2 MD3d n = 17, ipsi ctx vs contra ctx, paired t-test t = -1.64, P = 0.12; Nondep n= 10, t = 0.16, P = 0.88. Tet3 MD3d n = 17, ipsi ctx vs contra ctx, paired t-test t = -0.28, P 0.78; Nondep n = 12 t = -1.42, P = 0.18). (b) Apobec-1 and Apobec-3 mRNA expression was quantified by qPCR. The binocular area in the visual cortex of MD3d critical period mice was analyzed. Apobec-1 MD3d n = 10, ipsi ctx vs contra ctx, paired t-test t = -0.38, P = 0.71. Apobec-3 MD3d n = 10, ipsi ctx vs contra ctx, paired t-test t = -4.02, P = 0.002. Error bars represent SEM.
Post-translational regulation of TET proteins is under intense study14–16, moreover TET enzymatic activity was found to be highly regulated by cellular metabolites (e.g. alpha-ketoglutarate)17 and small molecules like ascorbic acid and L-cysteine18,19 suggesting that MD3d might influence the activity of TET or other factors involved in regulation of 5-hmC levels independently from a regulation of expression levels. These mechanisms, possibly together with enhanced Apobec-3 mediated deamination, might be involved in the decrease of 5-hmC observed
Supplementary Figure 10 Gene ontology analysis of RG108 regulated genes in the visual cortex of MD3d mice.
Singular enrichment analysis of KEGG pathway and biological processes. a) Upregulated genes, b) downregulated genes. The pie charts show some selected categories significantly enriched; the complete analysis is available in Supplementary Table 1.
Supplementary Figure 11 Gene ontology analysis of MD3d regulated genes in the visual cortex.
Singular enrichment analysis of KEGG pathway and biological processes. a) Upregulated genes, b) downregulated genes. The pie charts show some selected significantly enriched categories; the complete analysis is available in Supplementary Table 2.
Supplementary Figure 12 qPCR validation of RNA sequencing results.
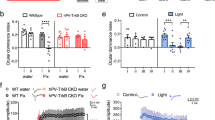
(a) Npas4 gene expression in the visual cortex of MD3d mice and MD3d mice treated with RG108 or vehicle. Monocular deprivation significantly downregulated the expression of Npas4 in the visual cortex contralateral to the deprived eye (cortex ipsilateral to the deprived eye vs. contralateral cortex n = 11, Paired t-test, t = -3.9, P = 0.002; MD3d vehicle vs MD3d RG108, n = 8, Student t-test, t = 4.43, P < 0.001). (b) Gadd45g decrease in the visual cortex contralateral to the deprived eye (data shown in Suppl. Fig. 3) was counteracted by RG108 infusion during the monocular deprivation (MD3d vehicle vs. MD3d RG108, n = 6, Student t-test, t = 2.48, P = 0.04). (c) qPCR in the visual cortex of MD3d critical period mice showed a significant downregulation of Nr4a1 in the contra ctx with respect to the ipsi ctx. (n=6, ipsi ctx vs contra ctx, paired t-test, t=3.07, P=0.02). The effect on Nr4a1 expression is completely reversed by RG108 treatment during MD3d (n=3, vehicle vs RG108, paired t-test, t=3.67, P=0.02). Error bars represent SEM.
Supplementary Figure 13 Sodium bisulfite sequencing in the visual cortex of MD3d critical period mice showed enhanced methylation on the CRE promoter region of Nr4a1.
Bisulfite sequencing of a CpG enriched region containing the CREB binding sequence in the Nr4a1 genomic locus in MD3d mice. This gene was selected for its upregulation by the DNMT inhibitor RG108 in our transcriptomic analysis. Average methylation (n=3 MD3d mice) was significantly increased by MD3d in the Contralateral cortex (contra ctx) (black squares) with respect to the ipsilateral cortex (ipsi ctx) (grey diamonds, paired t-test; p=0.012 t=2.93). Ten clones were sequenced for each cortex. Percentage of methylation of each CpG site represents the average of the percentage of clones showing cytosine methylation for each animal. Clones sequenced for the contra and ipsi ctx of a single representative mouse is reported under the graph. Black circles represent methylated cytosine.
Supplementary Figure 14 RG108 treatment does not change visual responses.
(a) Baseline activity of each unit of the different groups was not affected by RG108 or vehicle treatment (Nondep 77 units, MD3d untreated 77 units, MD3d RG108 71 units, MD3d vehicle 76 units. One way ANOVA F = 0.35, P = 0.79). (b) Responsiveness for each unit of the different groups (Nondep 77 units, MD3d untreated 77 units, MD3d RG108 71 units, MD3d vehicle 76 units) was expressed as a ratio between peak response frequency (measured on peristimulus time histograms as the maximum response of each unit) and baseline discharge frequency. No statistical difference is present between all groups (one way ANOVA F = 0.19, P = 0.91). (c) Receptive field size (RFS) was not altered by RG108 or vehicle infusion in MD3d critical period mice (Nondep 77 units, MD3d untreated 77 units, MD3d RG108 71 units, MD3d vehicle 76 units). None of the treatments significantly altered RFS (one way ANOVA F = 0.37, P = 0.77). Receptive fields were measured on peristimulus time histograms elicited by light bars of optimal orientation drifting in the visual field. The width of the part of visual field eliciting cell discharge greater than spontaneous discharge plus two SD was taken as RFS. Error bars represent SEM.
Supplementary information
Supplementary Text and Figures
Supplementary Tables 1–14 (PDF 923 kb)
Supplementary Table 1
MD3d RG108 RNA seq (XLSX 2823 kb)
Supplementary Table 2
MD3d RNA seq (XLSX 2846 kb)
Supplementary Table 3
Intersection (XLSX 16 kb)
Rights and permissions
About this article

Cite this article
Tognini, P., Napoli, D., Tola, J. et al. Experience-dependent DNA methylation regulates plasticity in the developing visual cortex. Nat Neurosci 18, 956–958 (2015). https://doi.org/10.1038/nn.4026
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nn.4026
This article is cited by
-
DNA Methylation-Dependent Gene Expression Regulation of Glutamate Transporters in Cultured Radial Glial Cells
Molecular Neurobiology (2022)
-
Retinal-input-induced epigenetic dynamics in the developing mouse dorsal lateral geniculate nucleus
Epigenetics & Chromatin (2019)
-
Mir-132/212 is required for maturation of binocular matching of orientation preference and depth perception
Nature Communications (2017)
-
Methyl-CpG binding domain proteins inhibit interspecies courtship and promote aggression in Drosophila
Scientific Reports (2017)
-
Bi-directional and shared epigenomic signatures following proton and 56Fe irradiation
Scientific Reports (2017)
