
Abstract
Receptor-interacting protein kinase 3 (RIPK3) functions as a key regulator of necroptosis. Here, we report that the RIPK3 expression level is negatively regulated by CHIP (carboxyl terminus of Hsp70-interacting protein; also known as STUB1) E3 ligase-mediated ubiquitylation. Chip−/− mouse embryonic fibroblasts and CHIP-depleted L929 and HT-29 cells exhibited higher levels of RIPK3 expression, resulting in increased sensitivity to necroptosis induced by TNF (also known as TNFα). These phenomena are due to the CHIP-mediated ubiquitylation of RIPK3, which leads to its lysosomal degradation. Interestingly, RIPK1 expression is also negatively regulated by CHIP-mediated ubiquitylation, validating the major role of CHIP in necrosome formation and sensitivity to TNF-mediated necroptosis. Chip−/− mice (C57BL/6) exhibit inflammation in the thymus and massive cell death and disintegration in the small intestinal tract, and die within a few weeks after birth. These phenotypes are rescued by crossing with Ripk3−/− mice. These results imply that CHIP is a bona fide negative regulator of the RIPK1–RIPK3 necrosome formation leading to desensitization of TNF-mediated necroptosis.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

References
Vanden Berghe, T., Linkermann, A., Jouan-Lanhouet, S., Walczak, H. & Vandenabeele, P. Regulated necrosis: the expanding network of non-apoptotic cell death pathways. Nat. Rev. Mol. Cell Biol. 15, 135–147 (2014).
Galluzzi, L. et al. Essential versus accessory aspects of cell death: recommendations of the NCCD 2015. Cell Death Differ. 22, 58–73 (2015).
Mocarski, E. S., Kaiser, W. J., Livingston-Rosanoff, D., Upton, J. W. & Daley-Bauer, L. P. True grit: programmed necrosis in antiviral host defense, inflammation, and immunogenicity. J. Immunol. 192, 2019–2026 (2014).
Linkermann, A. & Green, D. R. Necroptosis. N. Engl. J. Med. 370, 455–465 (2014).
Murphy, J. M. & Silke, J. Ars Moriendi; the art of dying well—new insights into the molecular pathways of necroptotic cell death. EMBO Rep. 15, 155–164 (2014).
Cho, Y. S. et al. Phosphorylation-driven assembly of the RIP1–RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell 137, 1112–1123 (2009).
He, S. et al. Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-α. Cell 137, 1100–1111 (2009).
Zhang, D. W. et al. RIP3, an energy metabolism regulator that switches TNF-induced cell death from apoptosis to necrosis. Science 325, 332–336 (2009).
Sun, L. et al. Mixed lineage kinase domain-like protein mediates necrosis signaling downstream of RIP3 kinase. Cell 148, 213–227 (2012).
Murphy, J. M. et al. The pseudokinase MLKL mediates necroptosis via a molecular switch mechanism. Immunity 39, 443–453 (2013).
Wu, J. et al. Mlkl knockout mice demonstrate the indispensable role of Mlkl in necroptosis. Cell Res. 23, 994–1006 (2013).
Remijsen, Q. et al. Depletion of RIPK3 or MLKL blocks TNF-driven necroptosis and switches towards a delayed RIPK1 kinase-dependent apoptosis. Cell Death Dis. 5, e1004 (2014).
Li, J. et al. The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis. Cell 150, 339–350 (2012).
Dondelinger, Y. et al. MLKL compromises plasma membrane integrity by binding to phosphatidylinositol phosphates. Cell Rep. 7, 971–981 (2014).
Wang, H. et al. Mixed lineage kinase domain-like protein MLKL causes necrotic membrane disruption upon phosphorylation by RIP3. Mol. Cell 54, 133–146 (2014).
Cai, Z. et al. Plasma membrane translocation of trimerized MLKL protein is required for TNF-induced necroptosis. Nat. Cell Biol. 16, 55–65 (2014).
Chen, X. et al. Translocation of mixed lineage kinase domain-like protein to plasma membrane leads to necrotic cell death. Cell Res. 24, 105–121 (2014).
Wu, X. N. et al. Distinct roles of RIP1–RIP3 hetero- and RIP3–RIP3 homo-interaction in mediating necroptosis. Cell Death Differ. 21, 1709–1720 (2014).
Cook, W. D. et al. RIPK1- and RIPK3-induced cell death mode is determined by target availability. Cell Death Differ. 21, 1600–1612 (2014).
Chen, W. et al. Ppm1b negatively regulates necroptosis through dephosphorylating Rip3. Nat. Cell Biol. 17, 434–444 (2015).
Ahmed, S. F. et al. The chaperone-assisted E3 ligase C terminus of Hsc70-interacting protein (CHIP) targets PTEN for proteasomal degradation. J. Biol. Chem. 287, 15996–16006 (2012).
Esser, C., Scheffner, M. & Hohfeld, J. The chaperone-associated ubiquitin ligase CHIP is able to target p53 for proteasomal degradation. J. Biol. Chem. 280, 27443–27448 (2005).
Ko, H. R. et al. P42 Ebp1 regulates the proteasomal degradation of the p85 regulatory subunit of PI3K by recruiting a chaperone-E3 ligase complex HSP70/CHIP. Cell Death Dis. 5, e1131 (2014).
Lee, E. W., Seo, J., Jeong, M., Lee, S. & Song, J. The roles of FADD in extrinsic apoptosis and necroptosis. BMB Rep. 45, 496–508 (2012).
Vanden Berghe, T., Kalai, M., van Loo, G., Declercq, W. & Vandenabeele, P. Disruption of HSP90 function reverts tumor necrosis factor-induced necrosis to apoptosis. J. Biol. Chem. 278, 5622–5629 (2003).
Park, S. Y., Shim, J. H. & Cho, Y. S. Distinctive roles of receptor-interacting protein kinases 1 and 3 in caspase-independent cell death of L929. Cell Biochem. Funct. 32, 62–69 (2014).
Petrucelli, L. et al. CHIP and Hsp70 regulate tau ubiquitination, degradation and aggregation. Hum. Mol. Genet. 13, 703–714 (2004).
Mollah, S. et al. Targeted mass spectrometric strategy for global mapping of ubiquitination on proteins. Rapid Commun. Mass Spectrom. 21, 3357–3364 (2007).
Kim, W. et al. Systematic and quantitative assessment of the ubiquitin-modified proteome. Mol. Cell 44, 325–340 (2011).
Sahara, N. et al. In vivo evidence of CHIP up-regulation attenuating tau aggregation. J. Neurochem. 94, 1254–1263 (2005).
Gunther, C. et al. Caspase-8 regulates TNF-α-induced epithelial necroptosis and terminal ileitis. Nature 477, 335–339 (2011).
Kaiser, W. J. et al. RIP3 mediates the embryonic lethality of caspase-8-deficient mice. Nature 471, 368–372 (2011).
Oberst, A. et al. Catalytic activity of the caspase-8-FLIP(L) complex inhibits RIPK3-dependent necrosis. Nature 471, 363–367 (2011).
Welz, P. S. et al. FADD prevents RIP3-mediated epithelial cell necrosis and chronic intestinal inflammation. Nature 477, 330–334 (2011).
Zhang, H. et al. Functional complementation between FADD and RIP1 in embryos and lymphocytes. Nature 471, 373–376 (2011).
Dillon, C. P. et al. RIPK1 blocks early postnatal lethality mediated by caspase-8 and RIPK3. Cell 157, 1189–1202 (2014).
Newton, K. et al. Activity of protein kinase RIPK3 determines whether cells die by necroptosis or apoptosis. Science 343, 1357–1360 (2014).
Rickard, J. A. et al. RIPK1 regulates RIPK3-MLKL-driven systemic inflammation and emergency hematopoiesis. Cell 157, 1175–1188 (2014).
Dannappel, M. et al. RIPK1 maintains epithelial homeostasis by inhibiting apoptosis and necroptosis. Nature 513, 90–94 (2014).
Takahashi, N. et al. RIPK1 ensures intestinal homeostasis by protecting the epithelium against apoptosis. Nature 513, 95–99 (2014).
Mandal, P. et al. RIP3 induces apoptosis independent of pronecrotic kinase activity. Mol. Cell 56, 481–495 (2014).
Orozco, S. et al. RIPK1 both positively and negatively regulates RIPK3 oligomerization and necroptosis. Cell Death Differ. 21, 1511–1521 (2014).
Duprez, L. et al. RIP kinase-dependent necrosis drives lethal systemic inflammatory response syndrome. Immunity 35, 908–918 (2011).
Onizawa, M. et al. The ubiquitin-modifying enzyme A20 restricts ubiquitination of the kinase RIPK3 and protects cells from necroptosis. Nat. Immunol. 16, 618–627 (2015).
Petersen, S. L. et al. TRAF2 is a biologically important necroptosis suppressor. Cell Death Differ. 22, 1846–1857 (2015).
Kang, T. B., Yang, S. H., Toth, B., Kovalenko, A. & Wallach, D. Caspase-8 blocks kinase RIPK3-mediated activation of the NLRP3 inflammasome. Immunity 38, 27–40 (2013).
Lawlor, K. E. et al. RIPK3 promotes cell death and NLRP3 inflammasome activation in the absence of MLKL. Nat. Commun. 6, 6282 (2015).
Vince, J. E. et al. Inhibitor of apoptosis proteins limit RIP3 kinase-dependent interleukin-1 activation. Immunity 36, 215–227 (2012).
Moriwaki, K. et al. The Necroptosis Adaptor RIPK3 promotes injury-induced cytokine expression and tissue repair. Immunity 41, 567–578 (2014).
Humphries, F., Yang, S., Wang, B. & Moynagh, P. N. RIP kinases: key decision makers in cell death and innate immunity. Cell Death Differ. 22, 225–236 (2015).
Silke, J., Rickard, J. A. & Gerlic, M. The diverse role of RIP kinases in necroptosis and inflammation. Nat. Immunol. 16, 689–697 (2015).
Min, J. N. et al. CHIP deficiency decreases longevity, with accelerated aging phenotypes accompanied by altered protein quality control. Mol. Cell. Biol. 28, 4018–4025 (2008).
Vanden Berghe, T. et al. Passenger mutations confound interpretation of all genetically modified congenic mice. Immunity 43, 200–209 (2015).
Schisler, J. C. et al. CHIP protects against cardiac pressure overload through regulation of AMPK. J. Clin. Invest. 123, 3588–3599 (2013).
Lee, I. H. et al. Ahnak functions as a tumor suppressor via modulation of TGFβ/Smad signaling pathway. Oncogene 33, 4675–4684 (2014).
Oh, W. et al. PML-IV functions as a negative regulator of telomerase by interacting with TERT. J Cell Sci. 122, 2613–2622 (2009).
Lee, E. W. et al. USP11-dependent selective cIAP2 deubiquitylation and stabilization determine sensitivity to Smac mimetics. Cell Death Differ. 22, 1463–1476 (2015).
Lee, E. W. et al. Differential regulation of p53 and p21 by MKRN1 E3 ligase controls cell cycle arrest and apoptosis. EMBO J. 28, 2100–2113 (2009).
Lee, E. W. et al. Ubiquitination and degradation of the FADD adaptor protein regulate death receptor-mediated apoptosis and necroptosis. Nat. Commun. 3, 978 (2012).
Acknowledgements
We thank S. Murata for the Chip−/− mice and V. M. Dixit for the Ripk3−/− mice. We thank the Cancer Metabolism Interest Group (cMIG) led by NCC Korea for discussion and advice. This research was supported by a grant from the National R&D Program for Cancer Control, Ministry of Health & Welfare, Republic of Korea (NCC-1420300) (to J.Song) and by a grant from the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2012R1A6A3A04040105) (to E.-W.L.) and by the Ministry of Science, ICT and Future Planning (NRF-2015R1A3A2066581) (to J.Song). Research in the Vandenabeele group is supported by Flemish grants (Research Foundation Flanders: FWO G.0875.11, FWO G.0973.11, FWO G.0A45.12N, FWO G.0787.13N, FWO G0E04.16N), a Methusalem grant (BOF16/MET_V/007), Ghent University grants (MRP, GROUP-ID consortium), a grant from the Foundation against Cancer (F94), and grants from VIB. Additionally, this research was partly supported by the BK21 Plus project of the National Research Foundation of Korea Grant (to J.Seo, M.J., H.-K.L., D.S., J.-H.K. and S.Y.H.) and by the grants for KMPC (Korea Mouse Phenotype Center) (to J.K.S.) from National Research Foundation of Korea (NRF), and C.L. acknowledges institutional support by KIST.
Author information
Authors and Affiliations
Contributions
J.Seo, E.-W.L. and J.Song conceived and designed the study. J.Seo, E.-W.L., D.S. and M.J. performed most of the experiments. J.Shin and C.L. performed the mass spectrometry analysis. J.Seo, H.S., D.S., H.-K.L., J.-H.K., S.Y.H. and J.K.S. performed the animal experiments and analysis. Y.D. and P.V. discussed the results, conceived some experiments, and provided critical reagents and comments. J.Seo, E.-W.L. and J.Song wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Ablation of CHIP increases RIPK3-mediated necroptosis in human HT-29 and mouse L929 cells.
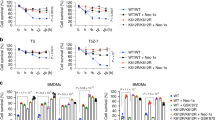
(a) HT-29 cells were transfected with the indicated siRNAs for 48 h and then treated with 30 ng/mL human TNFα (T), 5 μg/mL cycloheximide (C), and 30 μM z-VAD-fmk (Z) in the presence or absence of 30 μM Nec-1 (N) for 8 h. The cells were stained with Annexin V and 7-AAD, and then analysed by FACS. Representative plots of data are shown in left panel, and means ± S.D. from n = 3 independent experiments are presented in right panel. (b,c) HT-29 cells were transfected with siRNA against CHIP and/or RIPK3 and treated as in (a) for 10 h. Knockdown efficiency was confirmed by WB (b) and necrotic cells were detected by FACS (c). Representative plots of data are shown in left panel, and means ± S.D. from n = 3 independent experiments are presented in right panel (c). (d) L929 cells were transfected with the indicated siRNAs for 48 h and then treated with 5 ng/mL TNFα and 10 μM z-VAD-fmk in the presence or absence of Nec-1 for 2.5 h. The cells were stained with Annexin V and analysed by FACS. Representative plots of data from 3 independent experiments are shown in left panel and summarized in right panel. (e) L929 cells were transfected and treated as described in (d). Cell loss were determined using the Cell Titer Glo Luminescent Cell Viability Assay Kit. (f–h) L929 cells were transfected with the indicated siRNAs and treated with TNFα and z-VAD. The knockdown efficiencies of each siRNAs were shown by WB (f). Necrotic cells were determined by Annexin V staining and analysed by FACS (g). Cell loss was also determined as described above (h). (i,j) L929 cells stably expressing retroviral human CHIP (human CHIP shares all amino acids except for one amino acid with mouse CHIP) were transfected with the indicated siRNAs for 48 h. Cells were then treated with or without TNFα and z-VAD for 2.5 h. Expression of CHIP was analysed by WB (i). Cell death was determined by Annexin V staining (j). Data in (d,e,g,h,j) are means ± S, D.; n = 3 independent experiments (j). P values were determined by unpaired two-tailed Student’s t-test. (*P < 0.05, **P < 0.01, ***P < 0.001). Raw data from independent experiments are provided in Supplementary Table 1.
Supplementary Figure 2 CHIP does not affect TNF-mediated apoptosis.
(a) Primary Chip+/+ and Chip−/−MEFs were treated as indicated (T: mTNFα, 30 ng/mL; C: Cycloheximide, 2 μg/mL; Z: z-VAD-fmk, 30 μM; N: necrostatin-1, 30 μM) for 6 h or 12 h. The cells were stained with Annexin V and 7-AAD, and then analysed by FACS. Representative plots of data from n = 3 independent experiments are shown in left panel, and means ± S.D. are presented in right panel. (b) L929 cells were transfected with CHIP siRNA and treated as indicated for 2.5 h. Dead cells were determined by FACS analysis using Annexin V and 7-AAD staining. Representative plots of data from n = 3 independent experiments are shown in left panel, and means ± S.D. are presented in right panel. (c) HT-29 cells were transfected with CHIP siRNA and treated as indicated (T: hTNFα, 30 ng/mL; C: Cycloheximide, 5 μg/mL; Z: z-VAD-fmk, 30 μM; N: necrostatin-1, 30 μM) for 10 h or 12 h. The cells were stained with Annexin V and 7-AAD, and then analysed via FACS. Representative plots of data from n = 3 independent experiments are shown in left panel, and means ± S.D. are presented in right panel. P values were determined by unpaired two-tailed Student’s t-test. (*P < 0.05, **P < 0.01, ***P < 0.001). Raw data from independent experiments are provided in Supplementary Table 1.
Supplementary Figure 3 CHIP depletion facilitates necrosome formation by stabilising RIPK3 and RIPK1 proteins.
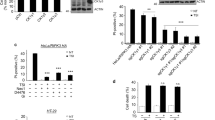
(a,b) HT-29 and L929 cells transfected with the siRNA against human CHIP or mouse CHIP for 48 h, respectively. HT-29 and L929 cells were then treated with hTNF, CHX, and zVAD (TCZ) or mTNF and CHX (TZ), respectively, as indicated. Cells were lysed with lysis buffer and immunoprecipitated with anti-Caspase-8 (a) or FADD (b). Associated proteins were detected by WB using the indicated antibodies. (c–e) HT-29 and L929 cells were transfected with control siRNA or siCHIP for 48 h. Expression of cell death related proteins were determined by WB in HT-29 and L929 cells with CHIP knockdown (c). Relative amounts of RIPK3 and RIPK1 were calculated and shown after normalising to Actin. mRNA levels were also measured by qRT-PCR (d,e). The data represent means ± S.D.; n = 3 independent experiments; **P < 0.01, ***P < 0.001 (d, e). (f) The stability of RIPK1 and RIPK3 in CHIP-depleted L929 cells was measured by treating the cells with cycloheximide for the indicated period, followed by WB analysis. Representative results of three independent experiments are shown. Relative amounts of RIPK3 and RIPK1 were calculated after normalising to Actin, and are shown in the graph. Half-lives of RIPK3 and RIPK1 were also calculated and presented in the graph.
Supplementary Figure 4 CHIP binds to the kinase and RHIM domains of RIPK3 and RIPK1, respectively.
(a,b) 293FT cells were transfected with plasmids expressing FLAG–CHIP and HA–RIPK3 or HA–RIPK1. The cell lysates were immunoprecipitated using an anti-FLAG or -HA antibodies, followed by WB using anti-RIPK3, -RIPK1 and -FLAG antibodies. (c,d) L929 cells were immunoprecipitated using an anti-RIPK3 (c) or –RIPK1 (d) antibody. Interactions between endogenous CHIP and RIPK3 or RIPK1 were then analysed by WB. (e,f) Bacterially produced GST and GST-CHIP were used for the in vitro binding analysis. HA-RIPK3 or HA-RIPK1 was produced using the TNT T7 reticulocyte system. GST or GST-CHIP was incubated with HA-RIPK3 or HA-RIPK1, and the sample was precipitated using Glutathione Sepharose beads. The interaction between CHIP and RIPKs was analysed by WB using anti-RIPK3 and anti-RIPK1 antibodies. The asterisk indicates N-terminal cleaved form of RIPK3 because anti-HA antibody did not detect the lower band. Only full-length RIPK3 bound to GST-CHIP, suggesting the N-terminal region of RIPK3 is required for the interaction with CHIP. (g,h) 293FT cells were transfected with the indicated plasmids. The cell lysates were immunoprecipitated using an anti-Myc antibody, and the interaction between these two proteins was analysed by WB. (i,j) 293FT cells were transfected with a combination of plasmids expressing of CHIP deficient mutants and RIPK3 or RIPK1. The cell lysates were immunoprecipitated using the indicated antibodies and then analysed via western blotting. (k,l) 293FT cells were transfected with a combination of CHIP wild type and RIPK3 or RIPK1 domain expressing plasmids and then immunoprecipitated using the indicated antibodies. The interaction between the peptides was analysed via western blotting using the indicated antibodies. (m) A schematic model of the interactions among RIPK1, RIPK3 and CHIP.
Supplementary Figure 5 CHIP is not involved in the geldanamycin-mediated degradation of RIPK3 or RIPK1 and induces the degradation of mutants RIPK3 and RIPK1 defective in aggresome formation.
(a) L929 cells were transfected with siCtrl or siCHIP and then treated with 1 μM geldanamycin for 16 h. The proteins were detected by WB using the indicated antibodies. (b,c) H1299 cells were co-transfected with plasmids expressing of FLAG-CHIP, HA-RIPK3 or HA-RIPK1 with GST, a transfection control, and then treated with geldanamycin for 12 h. The cells were analysed by WB. (d) H1299 cells were transfected with GFP-RIPK3 or GFP-RIPK1 in the absence or presence of FLAG-CHIP or FLAG-CHIP H260Q mutant and then treated with E64d + Pepstatin A for 16 h. The lysosomes and nucleus were stained using LysoTracker (red) and DAPI (blue). The cells were analysed via fluorescence microscopy. (e) H1299 cells were transfected with GFP-RIPK3 or GFP-RIPK1 in the absence or presence of WT or H260Q mutant of FLAG-CHIP and then treated with E64d-Pepstatin A. The cells were stained with an anti-FLAG antibody (red). (f) H1299 cells were transfected with GFP-RIPK3 or GFP-RIPK1 in the presence of FLAG-CHIP WT or mutant and then treated with E64d-Pepstatin A. Cells were stained with LysoTracker (red) and an anti-FLAG antibody (blue). (g) H1299 cells were transfected with the indicated plasmids and then treated with E64d-Pepstatin A. Cells were stained with LysoTracker (red) and an anti-FLAG antibody (blue). (h) H1299 cells were transfected with HA-RIPK3 V460P in the absence and presence of FLAG-CHIP, followed by WB using anti-HA, -FLAG and-GST antibodies. (i) H1299 cells were transfected with HA-RIPK1 I539D in the absence and presence of FLAG-CHIP, followed by WB using anti-HA, -FLAG and-GST antibodies. In d–g, a representative image is shown in the Fig. 5 and means from two independent experiments are presented in the graph. In each independent experiment, n = 100 cells were counted. All scale bars are 10 μm.
Supplementary Figure 6 Lysines of 55 and 363 of RIPK3 are responsible to CHIP depletion-induced RIPK3 up-regulation.
(a) 293FT cells were transfected with plasmids expressing the HA-RIPK3 WT or 4KR, K55⋅363R, K89⋅501R mutants in the absence or presence of Myc-CHIP. The lysates were immunoprecipitated using an anti-Myc antibody and then analysed via western blotting. (b) H1299 cells were transfected with HA-RIPK3 WT or K55⋅363R in the absence and presence of FLAG-CHIP, and treated with CHX as indicated. The proteins were detected by WB using anti-HA, -FLAG, -Actin antibodies, quantified by Image J, and are shown in the graph. Half-life of RIPK3 are presented in the graph. (c) H1299 cells were transfected with plasmids expressing GFP/RIPK3 K55⋅363R or K89⋅501R mutants in the absence or presence of FLAG-CHIP. Transfected cells were treated with E64d-Pepstatin A and LysoTracker for lysosome staining. Stained cells were analysed using fluorescence microscopy. A representative image is shown in the upper panel and means from two independent experiments are presented in the lower panel. In each independent experiment, n = 100 cells were counted. (d) Degradation of WT and single point mutant RIPK3 by CHIP was determined by WB in H1299 cells. (e,f) HeLa cells stably transfected with pBabe-RIPK3 WT, K55⋅363R and an empty vector were transfected with the indicated siRNAs for 48 h. Cells were treated with 10 ng/mL hTNFα, 5 μg/mL cycloheximide, and 20 μM z-VAD-fmk (TCZ) for 4 h (e). Cell loss was determined by Cell Titer Glo Luminescent Cell Viability Assay Kit (e), and protein levels were determined by western blot (f). Data are means ± S.D. n = 3 independent experiments (*P < 0.05, **P < 0.01). Raw data from independent experiments are provided in Supplementary Table 1. All scale bars are 10 μm.
Supplementary Figure 7 The K571⋅604⋅627R (3KR) RIPK1 are resistant to degradation and lysosomal localisation by CHIP.
(a) H1299 cells were transfected with HA-RIPK1 mutants with or without FLAG-CHIP as indicated. Degradation of each mutant by CHIP were determined by WB. (b) The amino acid sequence alignment of human and mouse RIPK1. (c) 293FT cells were transfected with plasmids expressing the HA-RIPK1 WT or 3KR mutant in the absence or presence of Myc-CHIP. The lysates were immunoprecipitated using an anti-Myc antibody and then analysed via western blotting. (d) 293FT cells transfected with the plasmid expression HA-RIPK1 (WT and 3KR) and His-Ub with or without FLAG-CHIP in the presence and absence of E64d-Pepstatin A. A pull-down assay was performed using Ni2+-NTA beads. The level of RIPK1 ubiquitylation was determined by WB using an anti-RIPK1 antibody. (e) H1299 cells were transfected with a combination of GFP/RIPK1 WT or 3KR and FLAG-CHIP and were treated with E64d/Pepstatin A and LysoTracker. The transfected cells were stained using an anti-FLAG antibody and then analysed via fluorescence microscopy. A representative image is shown in the left panel and means from two independent experiments are presented in the right panel. In each independent experiment, n = 100 cells were counted. All scale bars are 10 μm.
Supplementary Figure 8 The analysis of Chip−/− and Chip−/− Ripk3−/− mice in spleen, thymus and small intestine.
(a) Genomic DNA was prepared via phenol-extraction. The genotypes were determined via PCR using specific primers for each gene. (b) The expected and observed frequencies and the ratio of each genotype in the littermates from the crosses of Chip+/− x Chip+/− mice and Chip+/−Ripk3−/− x Chip+/−Ripk3−/− mice. (c) Representative spleen pictures and weight of WT (n = 13; male = 4, female = 9), Chip−/− (n = 8; male = 3, female = 5) and DKO (n = 9; male = 3, female = 6) at 4 weeks of age. Data are means ± S.D.; n = number of mice analysed; *P < 0.05. Raw data are presented in Supplementary Table 1. (d) Representative pictures of WT (n = 3; female = 3) and DKO (n = 3; female = 3) thymus and spleen at 16 weeks of age. n means number of mice analysed. (e) Representative H&E-stained of sections of WT and DKO small intestine at 16 weeks of age (n = 3 for each genotype). Scale bars, 100 μm.
Supplementary information
Supplementary Information
Supplementary Information (PDF 18363 kb)
Supplementary Table 1
Supplementary Information (XLSX 45 kb)
Rights and permissions
About this article

Cite this article
Seo, J., Lee, EW., Sung, H. et al. CHIP controls necroptosis through ubiquitylation- and lysosome-dependent degradation of RIPK3. Nat Cell Biol 18, 291–302 (2016). https://doi.org/10.1038/ncb3314
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3314
This article is cited by
-
MLKL polymerization-induced lysosomal membrane permeabilization promotes necroptosis
Cell Death & Differentiation (2024)
-
Mediators of necroptosis: from cell death to metabolic regulation
EMBO Molecular Medicine (2024)
-
The therapeutic potential of targeting regulated non-apoptotic cell death
Nature Reviews Drug Discovery (2023)
-
RIPK3 promoter hypermethylation in hepatocytes protects from bile acid-induced inflammation and necroptosis
Cell Death & Disease (2023)
-
PRMT5-mediated regulatory arginine methylation of RIPK3
Cell Death Discovery (2023)
