
Abstract
During meiosis, crossover recombination connects homologous chromosomes to direct their accurate segregation1. Defective crossing over causes infertility, miscarriage and congenital disease. Each pair of chromosomes attains at least one crossover via the formation and biased resolution of recombination intermediates known as double Holliday junctions2,3. A central principle of crossover resolution is that the two Holliday junctions are resolved in opposite planes by targeting nuclease incisions to specific DNA strands4. The endonuclease activity of the MutLγ complex has been implicated in the resolution of crossovers5,6,7,8,9,10, but the mechanisms that activate and direct strand-specific cleavage remain unknown. Here we show that the sliding clamp PCNA is important for crossover-biased resolution. In vitro assays with human enzymes show that PCNA and its loader RFC are sufficient to activate the MutLγ endonuclease. MutLγ is further stimulated by a co-dependent activity of the pro-crossover factors EXO1 and MutSγ, the latter of which binds Holliday junctions11. MutLγ also binds various branched DNAs, including Holliday junctions, but does not show canonical resolvase activity, implying that the endonuclease incises adjacent to junction branch points to achieve resolution. In vivo, RFC facilitates MutLγ-dependent crossing over in budding yeast. Furthermore, PCNA localizes to prospective crossover sites along synapsed chromosomes. These data highlight similarities between crossover resolution and the initiation steps of DNA mismatch repair12,13 and evoke a novel model for crossover-specific resolution of double Holliday junctions during meiosis.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


Data availability
Relevant data generated or analysed during this study are included in this Article and its Supplementary Information files. Biological materials are available from the corresponding author.
Change history
19 January 2021
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Hunter, N. Meiotic recombination: the essence of heredity. Cold Spring Harb. Perspect. Biol. 7, a016618 (2015).
Allers, T. & Lichten, M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106, 47–57 (2001).
Marsolier-Kergoat, M. C., Khan, M. M., Schott, J., Zhu, X. & Llorente, B. Mechanistic view and genetic control of DNA recombination during meiosis. Mol. Cell 70, 9–20.e6 (2018).
Szostak, J. W., Orr-Weaver, T. L., Rothstein, R. J. & Stahl, F. W. The double-strand-break repair model for recombination. Cell 33, 25–35 (1983).
Nishant, K. T., Plys, A. J. & Alani, E. A mutation in the putative MLH3 endonuclease domain confers a defect in both mismatch repair and meiosis in Saccharomyces cerevisiae. Genetics 179, 747–755 (2008).
Zakharyevich, K., Tang, S., Ma, Y. & Hunter, N. Delineation of joint molecule resolution pathways in meiosis identifies a crossover-specific resolvase. Cell 149, 334–347 (2012).
Toledo, M. et al. A mutation in the endonuclease domain of mouse MLH3 reveals novel roles for MutLγ during crossover formation in meiotic prophase I. PLoS Genet. 15, e1008177 (2019).
Ranjha, L., Anand, R. & Cejka, P. The Saccharomyces cerevisiae Mlh1-Mlh3 heterodimer is an endonuclease that preferentially binds to Holliday junctions. J. Biol. Chem. 289, 5674–5686 (2014).
Rogacheva, M. V. et al. Mlh1-Mlh3, a meiotic crossover and DNA mismatch repair factor, is a Msh2-Msh3-stimulated endonuclease. J. Biol. Chem. 289, 5664–5673 (2014).
Manhart, C. M. et al. The mismatch repair and meiotic recombination endonuclease Mlh1-Mlh3 is activated by polymer formation and can cleave DNA substrates in trans. PLoS Biol. 15, e2001164 (2017).
Snowden, T., Acharya, S., Butz, C., Berardini, M. & Fishel, R. hMSH4-hMSH5 recognizes Holliday Junctions and forms a meiosis-specific sliding clamp that embraces homologous chromosomes. Mol. Cell 15, 437–451 (2004).
Iyer, R. R., Pluciennik, A., Burdett, V. & Modrich, P. L. DNA mismatch repair: functions and mechanisms. Chem. Rev. 106, 302–323 (2006).
Kolodner, R. D. A personal historical view of DNA mismatch repair with an emphasis on eukaryotic DNA mismatch repair. DNA Repair (Amst.) 38, 3–13 (2016).
Manhart, C. M. & Alani, E. Roles for mismatch repair family proteins in promoting meiotic crossing over. DNA Repair (Amst.) 38, 84–93 (2016).
Kadyrova, L. Y. & Kadyrov, F. A. Endonuclease activities of MutLα and its homologs in DNA mismatch repair. DNA Repair (Amst.) 38, 42–49 (2016).
Kadyrov, F. A., Dzantiev, L., Constantin, N. & Modrich, P. Endonucleolytic function of MutLα in human mismatch repair. Cell 126, 297–308 (2006).
Pluciennik, A. et al. PCNA function in the activation and strand direction of MutLα endonuclease in mismatch repair. Proc. Natl Acad. Sci. USA 107, 16066–16071 (2010).
Pillon, M. C. et al. Structure of the endonuclease domain of MutL: unlicensed to cut. Mol. Cell 39, 145–151 (2010).
Pillon, M. C. et al. The sliding clamp tethers the endonuclease domain of MutL to DNA. Nucleic Acids Res. 43, 10746–10759 (2015).
Claeys Bouuaert, C. & Keeney, S. Distinct DNA-binding surfaces in the ATPase and linker domains of MutLγ determine its substrate specificities and exert separable functions in meiotic recombination and mismatch repair. PLoS Genet. 13, e1006722 (2017).
Kadyrova, L. Y., Gujar, V., Burdett, V., Modrich, P. L. & Kadyrov, F. A. Human MutLγ, the MLH1-MLH3 heterodimer, is an endonuclease that promotes DNA expansion. Proc. Natl Acad. Sci. USA 117, 3535–3542 (2020).
Kadyrov, F. A. et al. Saccharomyces cerevisiae MutLα is a mismatch repair endonuclease. J. Biol. Chem. 282, 37181–37190 (2007).
Sonntag Brown, M., Lim, E., Chen, C., Nishant, K. T. & Alani, E. Genetic analysis of mlh3 mutations reveals interactions between crossover promoting factors during meiosis in baker’s yeast. G3 (Bethesda) 3, 9–22 (2013).
Lahiri, S., Li, Y., Hingorani, M. M. & Mukerji, I. MutSγ-induced dna conformational changes provide insights into its role in meiotic recombination. Biophys. J. 115, 2087–2101 (2018).
Gray, S. & Cohen, P. E. Control of meiotic crossovers: from double-strand break formation to designation. Annu. Rev. Genet. 50, 175–210 (2016).
Zakharyevich, K. et al. Temporally and biochemically distinct activities of Exo1 during meiosis: double-strand break resection and resolution of double Holliday junctions. Mol. Cell 40, 1001–1015 (2010).
Schaetzlein, S. et al. Mammalian Exo1 encodes both structural and catalytic functions that play distinct roles in essential biological processes. Proc. Natl Acad. Sci. USA 110, E2470–E2479 (2013).
Mukherjee, S., Wright, W. D., Ehmsen, K. T. & Heyer, W. D. The Mus81-Mms4 structure-selective endonuclease requires nicked DNA junctions to undergo conformational changes and bend its DNA substrates for cleavage. Nucleic Acids Res. 42, 6511–6522 (2014).
Sisakova, A., Altmannova, V., Sebesta, M. & Krejci, L. Role of PCNA and RFC in promoting Mus81-complex activity. BMC Biol. 15, 90 (2017).
Zhang, L. et al. Topoisomerase II mediates meiotic crossover interference. Nature 511, 551–556 (2014).
Genschel, J., Bazemore, L. R. & Modrich, P. Human exonuclease I is required for 5′ and 3′ mismatch repair. J. Biol. Chem. 277, 13302–13311 (2002).
Eichman, B. F., Vargason, J. M., Mooers, B. H. & Ho, P. S. The Holliday junction in an inverted repeat DNA sequence: sequence effects on the structure of four-way junctions. Proc. Natl Acad. Sci. USA 97, 3971–3976 (2000).
Kolas, N. K. et al. Localization of MMR proteins on meiotic chromosomes in mice indicates distinct functions during prophase I. J. Cell Biol. 171, 447–458 (2005).
Nishimura, K., Fukagawa, T., Takisawa, H., Kakimoto, T. & Kanemaki, M. An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nat. Methods 6, 917–922 (2009).
Tang, S., Wu, M. K. Y., Zhang, R. & Hunter, N. Pervasive and essential roles of the Top3-Rmi1 decatenase orchestrate recombination and facilitate chromosome segregation in meiosis. Mol. Cell 57, 607–621 (2015).
Morawska, M. & Ulrich, H. D. An expanded tool kit for the auxin-inducible degron system in budding yeast. Yeast 30, 341–351 (2013).
Benjamin, K. R., Zhang, C., Shokat, K. M. & Herskowitz, I. Control of landmark events in meiosis by the CDK Cdc28 and the meiosis-specific kinase Ime2. Genes Dev. 17, 1524–1539 (2003).
Carlile, T. M. & Amon, A. Meiosis I is established through division-specific translational control of a cyclin. Cell 133, 280–291 (2008).
Louvion, J. F., Havaux-Copf, B. & Picard, D. Fusion of GAL4-VP16 to a steroid-binding domain provides a tool for gratuitous induction of galactose-responsive genes in yeast. Gene 131, 129–134 (1993).
Owens, S., Tang, S. & Hunter, N. Monitoring recombination during meiosis in budding yeast. Methods Enzymol. 601, 275–307 (2018).
Hunter, N. & Kleckner, N. The single-end invasion: an asymmetric intermediate at the double-strand break to double-Holliday junction transition of meiotic recombination. Cell 106, 59–70 (2001).
Grubb, J., Brown, M. S. & Bishop, D. K. Surface spreading and immunostaining of yeast chromosomes. J. Vis. Exp. 102, e53081 (2015).
Dzantiev, L. et al. A defined human system that supports bidirectional mismatch-provoked excision. Mol. Cell 15, 31–41 (2004).
Cai, J. et al. Reconstitution of human replication factor C from its five subunits in baculovirus-infected insect cells. Proc. Natl Acad. Sci. USA 93, 12896–12901 (1996).
Acknowledgements
We thank P. Modrich, J. Hurwitz, K. Marians, S. Kowalczykowski, P. Cohen, S. Keeney and A. Shinohara for reagents; P. Cejka for reagents and communicating unpublished data; B. Llorente and members of the Hunter laboratory for support and discussions. This work was supported by NIH NIGMS grant GM074223 to N.H. S.N.O. was supported by NIH NIGMS T32 Training Program in Molecular and Cellular Biology 5T32GM007377 and an F31 Ruth L. Kirschstein National Research Service Award 1F31GM125106. M.I. was supported by a Japan Society for the Promotion of Science postdoctoral fellowship for research abroad. N.H. is an Investigator of the Howard Hughes Medical Institute.
Author information
Authors and Affiliations
Contributions
D.S.K., S.N.O., M.H. and N.H. conceived the study and designed most experiments. D.S.K. and M.H. performed all enzyme purifications and assays. S.N.O. performed all yeast experiments. M.I. performed the mouse cytology. All other authors assisted with experiments and data analysis. D.S.K., S.N.O. and N.H. wrote the manuscript with inputs and edits from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Andreas Hochwagen and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
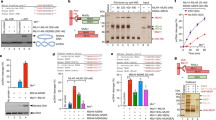
Extended Data Fig. 1 Human MutLγ purification and endonuclease activity.
a, Human MLH1 and MLH3 expression constructs highlighting domain structure and affinity-purification tags: MLH3 ID, human MLH3 interaction domain; endo, endonuclease domain; MBP, maltose binding protein; PP, PreScission protease cleavage site; 10His, deca-histidine tag. Sequences of the conserved metal binding site for (1) wild-type and (2) the nuclease-dead D1223N mutant of MLH3 are also shown. b, Representative MutLγ purification steps monitored by 10% SDS–PAGE stained with Coomassie blue. Amylose enriched fractions were treated with PreScission Protease, to cleave the maltose-binding protein tag, and then subjected to Ni-NTA affinity purification. c, d, Endonuclease assays for varying concentrations of MutLγ and MutLγ(D1223N) (0–300 nM protein with 5 mM Mn2+ (c) or Mg2+ (d)) incubated at 37 °C for 60 min. e, Quantitation of experiments represented in c and d. Mean ± s.e.m. are shown for three experiments after subtracting background nicked product from no-protein controls.
Extended Data Fig. 2 MutLγ endonuclease activity with various metal cofactors and stimulation by ATP.
a, Representative gel image showing MutLγ endonuclease assays with various metal ions (100 nM MutLγ and 5 mM indicated divalent metal ions, incubated at 37 °C for 90 min). Migration positions of supercoiled (S) plasmid and nicked (N) product are shown. For reference, plasmid linearized (L) with BamHI is also shown. Mean ± s.e.m. are shown for three experiments after subtracting background nicked product from no-protein controls. b, Representative endonuclease assays with 100 nM MutLγ in 5 mM Mg2+ or Mn2+ with the addition of various second metal cofactors (5 mM). Reactions were incubated at 37 °C for 90 min. Metals other than Mg2+ compete with Mn2+ to inhibit MutLγ endonuclease activity. Mean ± s.e.m. are shown for three experiments. c, Representative gel image and quantification of human MutLγ endonuclease activity with and without ATP and metal cofactors (100 nM MutLγ, 0.5 mM ATP and 5 mM Mn2+ and/or Mg2+ incubated at 37 °C for 60 min). Mean ± s.e.m. are shown for five experiments. d, MutLγ endonuclease activity with increasing ATP concentration. 100 nM MutLγ was incubated with 5 mM Mn2+ and Mg2+ and indicated concentrations of ATP at 37 °C for 60 min. Endonuclease stimulation was optimal with 0.5 mM ATP, while higher concentrations were inhibitory. Mean ± s.e.m. are shown for three independent experiments.
Extended Data Fig. 3 Purification and characterization of human MutSγ.
a, Representative MutSγ purification steps monitored by 10% SDS–PAGE stained with Coomassie blue. b, Representative images of EMSAs analysing MutSγ binding to 80-mer single and double-stranded DNAs. Binding reactions contained 1 nM 32P-labelled DNA, 100 mM NaCl and 5 mM Mg2+ and were incubated on ice for 10 min. Bound and free DNA species were resolved by non-denaturing 5% polyacrylamide gel electrophoresis and processed for imaging. c, Representative images of EMSAs analysing MutSγ binding to pro-HJ and Holliday junction structures. Terminally 32P-labelled strands are indicated by asterisks. d, Quantification of the EMSAs represented in b and c showing mean ± s.e.m. for three independent experiments. e, SDS–PAGE analysis of purified nuclease-dead human EXO1(D173A) (10% gel stained with Coomassie). f, Endonuclease assays with varying mixtures of MutLγ, MutSγ and EXO1(D173A), with and without ATP (50 nM each protein, 0.5 mM ATP, 5 mM Mn2+ and Mg2+ incubated at 37 °C for 60 min). Mean ± s.e.m. are shown for three experiments. g, Negative control endonuclease assays with nuclease-dead human MutLγ(D1223N), MutSγ and EXO1(D173A). Average percentage nicking ± s.e.m. are shown for three experiments.
Extended Data Fig. 4 Colocalization of MutLγ and MutSγ in mouse spermatocytes.
a, Representative images of surface-spread spermatocyte chromosomes from the indicated pachytene substages, immuno-stained for MSH4 (green), MLH1 (magenta) and the chromosome axis marker SYCP3 (blue). Magnified panels show individual pairs of synapsed chromosomes. Scale bars, 10 and 2 μm, respectively. Arrowheads indicate crossover-specific MLH1 foci that colocalize with MSH4. b, Quantification of MSH4 and MLH1 foci and MSH4–MLH1 cofoci in early, mid and late-pachytene stages (n = 12, 15 and 13 nuclei, respectively). Error bars indicate mean ± s.e.m. N/A, not applicable because MLH1 foci were not observed in early pachytene. c, Percentage of MSH4 foci that are colocalized with MLH1 in mid and late pachytene nuclei (n = 12, 15 and 13 nuclei, respectively). Error bars indicate mean ± s.e.m. The percentage of co-foci increases as most MSH4 foci disappear in late pachynema.
Extended Data Fig. 5 RFC–PCNA activates the latent endonuclease activity of MutLγ in Mg2+.
a, b, SDS–PAGE analysis of purified PCNA and RFC (10% gels stained with Coomassie). c, d, Time-course analysis of MutLγ endonuclease activity in Mg2+, with and without RFC–PCNA and MutSγ–EXO1(D173A). Representative gel images (c) and quantification of three independent experiments (d, mean ± s.e.m.) are shown. Reactions contained 25 nM indicated proteins, 0.5 mM ATP and 5 mM Mg2+ and were incubated at 37 °C for the indicated times. e, Experimental scheme to analyse MutLγ–PCNA interaction in solution. f, SYPRO-Ruby stained SDS–PAGE analysis of MutLγ–PCNA interaction. g, ATP dependence of ensemble nicking reactions showing a representative gel image and quantification of percentage nicking (mean ± s.e.m. from three independent experiments). Reaction conditions were as described for c and d but without ATP, except in the positive control reaction. h, Dependence of ensemble nicking reactions on the MLH3 endonuclease active site. Representative gel images for reactions with nuclease-dead MutLγ(D1223N) and quantification of the percentage nicking are shown (mean ± s.e.m. from three independent experiments). Reactions contained 25 nM indicated proteins, 0.5 mM ATP and 5 mM Mn2+ and were incubated at 37 °C for 30 min. A reaction containing wild-type human MutLγ was included as a positive control.
Extended Data Fig. 6 Human MutLγ binding to single-stranded DNA.
a, Representative image of EMSAs comparing human MutLγ binding to ssDNA and Holliday junctions. b, Quantification of the EMSAs represented in a. Mean ± s.e.m. for three independent experiments are shown. Binding reactions contained 1 nM 32P-labelled DNA, 100 mM NaCl and 5 mM Mg2+ and were incubated on ice for 10 min. Bound and free DNA species were resolved by non-denaturing 5% polyacrylamide gel electrophoresis and processed for imaging. c, Representative images of EMSAs analysing human MutLγ binding to 80-mer DNAs containing single-strand gaps of varying size. To define the minimal single-strand gap that is efficiently bound by MutLγ, 80-nt linear DNAs containing gaps of varying lengths were tested. d, Quantification of the EMSAs represented in c showing mean ± s.e.m. for three independent experiments. Significant binding was not detected for gaps of 5 and 10 nt, but 20 and 40 nt gaps were bound efficiently. Notably, this minimal binding site is similar to that bound by the canonical ssDNA binding protein RPA. e, Representative images of EMSAs for MutLγ binding to an 80mer DNA containing a 40-nucleotide gap, with or without previous incubation with RPA. f, Quantification of the EMSAs represented in e shows that human MutLγ binds a 40 nt gap with almost identical affinity as a 40-mer single-stranded DNA; and that pre-incubation with RPA blocks MutLγ binding. Error bars show mean ± s.e.m. for three independent experiments. g, Representative images of EMSAs for MutLγ binding to pro-HJs and gapped-HJs that were pre-incubated with RPA. To test whether the branched and ssDNA binding activities are autonomous, gHJ and pHJ substrates were pre-incubated with RPA and then used as binding substrates for MutLγ. RPA-gHJ and RPA–pHJ complexes were still readily bound by MutLγ. h, Quantification of the EMSAs represented in g and in Fig. 3a shows that ssDNA and junction binding activities of MutLγ are largely independent. Error bars show mean ± s.e.m. for three independent experiments.
Extended Data Fig. 7 Model Holliday junctions are not incised by MutLγ.
a, b, Representative gel images of ensemble endonuclease reactions with Holliday junction (HJ) and nicked Holliday junction (nHJ) substrates. Reactions contained 25 nM indicated proteins, 0.5 mM ATP and 5 mM Mg2+/Mn2+, and were incubated at 37 °C for 30 min. The reaction products were analysed on a 15% denaturing polyacrylamide gel to detect strand nicking at any position. Nicking was not detected consistent with non-denaturing gels showing that model Holliday junction and nicked Holliday junction structures are not resolved by the MutLγ ensemble (not shown).
Extended Data Fig. 8 Cleavage of supercoiled plasmid containing a cruciform structure.
a, Endonuclease assays with pUC(AT) and varying mixtures of human MutLγ, MutSγ and EXO1(D173A), with and without ATP (50 nM each protein, 0.5 mM ATP, 5 mM Mn2+ and 5 mM Mg2+, incubated at 37 °C for 60 min). Mean ± s.e.m. are shown for three experiments. Under these conditions, pUC(AT) is incised 1.3–1.7-fold more efficiently than the corresponding reactions with pUC19 (also see Fig. 3e). Notably, the linear cleavage product seen with the pUC19 substrate (expected position indicated by L*; see Fig. 1e) was not observed. We suggest that this reflects a more physiological nicking reaction because MutSγ and MutLγ can interact with the cruciform structure. Thus, both the efficiency of nicking and the nature of the incision products are altered when a Holliday junction is present. b, The interdependent nature of stimulation by MutSγ and EXO1(D173A). Representative gel image and quantification of ensemble endonuclease reactions with using pUC(AT) as substrate and Mg2+ as the sole metal cofactor. Reactions contained 25 nM indicated proteins, 0.5 mM ATP and 5 mM Mg2+ and were incubated at 37 °C for 30 min. Mean ± s.e.m. are shown for three experiments. c, d, Negative control ensemble endonuclease assays for pUC(AT) (b) and pUC19 (c) containing nuclease-dead MutLγ(D1223N). Reactions contained 25 nM indicated proteins, 0.5 mM ATP and 5 mM Mg2+ or Mn2+ and were incubated at 37 °C for 30 min. Average percent nicking ± s.e.m. is shown for three independent assays.
Extended Data Fig. 9 Physical assay system for analysis of meiotic recombination in budding yeast.
a, Map of the HIS4:LEU2 recombination hotspot locus highlighting positions of the initiating DSB site, XhoI restriction sites (circled Xs) and the probe used in Southern blotting. Sizes of diagnostic fragments are shown to the right. b, Representative 1D gel Southern blot images for analysis of crossovers and dHJs in RFC1-AID and RFC1-AID mlh3∆ strains, with and without the addition of auxin to trigger Rfc1-AID degradation. Time points are 0, 7, 8, 9 and 11 h after induction of meiosis.
Extended Data Fig. 10 Model variants.
Comparison of variations on the basic theme that MutLγ mediates crossover-specific dHJ resolution by catalysing strand-specific incisions on both sides of the two HJs. a, The model presented in Fig. 4h is shown for comparison. This version has recombination-associated DNA synthesis tracts terminating outside the dHJ. PCNA at these termini direct the MutLγ–MutSγ–Exo1 ensemble to incise upstream (5′) specifically on the strands with DNA synthesis, maintaining this specificity across the stacked junctions (lower panels). Resolution of HJ-1 results in a crossover whereas HJ-2 is resolved as a non-crossover. Notably, this version of the model produces only ‘pattern 1’ crossovers, shown previously to be promoted by MutLγ3. b, By contrast, a more canonical scheme, with synthesis tracts and associated PCNA molecules located within the dHJ produces ‘pattern 2’ crossovers, thereby disfavouring this version of the model. c, Pattern 1 crossovers are also produced by a variant of the model in which MutLγ–MutSγ–Exo1 incises downstream (3′) of the internal synthesis tracts. However, this polarity is opposite to that established for MMR12,13.
Supplementary information
Supplementary Figure
This file contains the uncropped gel images.
Supplementary Tables
This file contains a list of oligonucleotides used in the study (Supplementary Table 1), and a list of strains used in the study (Supplementary Table 2).
Rights and permissions
About this article

Cite this article
Kulkarni, D.S., Owens, S.N., Honda, M. et al. PCNA activates the MutLγ endonuclease to promote meiotic crossing over. Nature 586, 623–627 (2020). https://doi.org/10.1038/s41586-020-2645-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2645-6
This article is cited by
-
Seeding the meiotic DNA break machinery and initiating recombination on chromosome axes
Nature Communications (2024)
-
Divergence and conservation of the meiotic recombination machinery
Nature Reviews Genetics (2023)
-
Gene expression profiling and protein–protein interaction analysis reveals the dynamic role of MCM7 in Alzheimer's disorder and breast cancer
3 Biotech (2022)
-
Concerted cutting by Spo11 illuminates meiotic DNA break mechanics
Nature (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
