
Abstract
Large-scale genomic studies have shown that half of epithelial ovarian cancers (EOCs) have alterations in genes regulating homologous recombination (HR) repair1. Loss of HR accounts for the genomic instability of EOCs and for their cellular hyper-dependence on alternative poly-ADP ribose polymerase (PARP)-mediated DNA repair mechanisms2,3,4,5. Previous studies have implicated the DNA polymerase θ (Polθ also known as POLQ, encoded by POLQ)6 in a pathway required for the repair of DNA double-strand breaks7,8,9, referred to as the error-prone microhomology-mediated end-joining (MMEJ) pathway10,11,12,13. Whether Polθ interacts with canonical DNA repair pathways to prevent genomic instability remains unknown. Here we report an inverse correlation between HR activity and Polθ expression in EOCs. Knockdown of Polθ in HR-proficient cells upregulates HR activity and RAD51 nucleofilament assembly, while knockdown of Polθ in HR-deficient EOCs enhances cell death. Consistent with these results, genetic inactivation of an HR gene (Fancd2) and Polq in mice results in embryonic lethality. Moreover, Polθ contains RAD51 binding motifs and it blocks RAD51-mediated recombination. Our results reveal a synthetic lethal relationship between the HR pathway and Polθ-mediated repair in EOCs, and identify Polθ as a novel druggable target for cancer therapy.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
References
The Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 474, 609–615 (2011)
Farmer, H. et al. Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature 434, 917–921 (2005)
Bryant, H. E. et al. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature 434, 913–917 (2005)
Kennedy, R. D. & D’Andrea, A. D. DNA repair pathways in clinical practice: lessons from pediatric cancer susceptibility syndromes. J. Clin. Oncol. 24, 3799–3808 (2006)
Bast, R. C., Jr, Hennessy, B. & Mills, G. B. The biology of ovarian cancer: new opportunities for translation. Nature Rev. Cancer 9, 415–428 (2009)
Yousefzadeh, M. J. & Wood, R. D. DNA polymerase POLQ and cellular defense against DNA damage. DNA Repair 12, 1–9 (2013)
Shima, N., Munroe, R. J. & Schimenti, J. C. The mouse genomic instability mutation chaos1 is an allele of Polq that exhibits genetic interaction with Atm. Mol. Cell. Biol. 24, 10381–10389 (2004)
Yoshimura, M. et al. Vertebrate POLQ and POLβ cooperate in base excision repair of oxidative DNA damage. Mol. Cell 24, 115–125 (2006)
Muzzini, D. M., Plevani, P., Boulton, S. J., Cassata, G. & Marini, F. Caenorhabditis elegans POLQ-1 and HEL-308 function in two distinct DNA interstrand cross-link repair pathways. DNA Repair 7, 941–950 (2008)
McVey, M. & Lee, S. E. MMEJ repair of double-strand breaks (director’s cut): deleted sequences and alternative endings. Trends in Genet. 24, 529–538 (2008)
Chan, S. H., Yu, A. M. & McVey, M. Dual roles for DNA polymerase theta in alternative end-joining repair of double-strand breaks in Drosophila. PLoS Genet. 6, e1001005 (2010)
Yu, A. M. & McVey, M. Synthesis-dependent microhomology-mediated end joining accounts for multiple types of repair junctions. Nucleic Acids Res. 38, 5706–5717 (2010)
Koole, W. et al. A polymerase theta-dependent repair pathway suppresses extensive genomic instability at endogenous G4 DNA sites. Nat. Commun. 5, 3216 (2014)
Nakanishi, K. et al. Human Fanconi anemia monoubiquitination pathway promotes homologous DNA repair. Proc. Natl Acad. Sci. USA 102, 1110–1115 (2005)
Moldovan, G. L. et al. Inhibition of homologous recombination by the PCNA-interacting protein PARI. Mol. Cell 45, 75–86 (2012)
Ira, G., Malkova, A., Liberi, G., Foiani, M. & Haber, J. E. Srs2 and Sgs1-Top3 suppress crossovers during double-strand break repair in yeast. Cell 115, 401–411 (2003)
Ward, J. D. et al. Overlapping mechanisms promote postsynaptic RAD-51 filament disassembly during meiotic double-strand break repair. Mol. Cell 37, 259–272 (2010)
Seki, M., Marini, F. & Wood, R. D. POLQ (Pol theta), a DNA polymerase and DNA-dependent ATPase in human cells. Nucleic Acids Res. 31, 6117–6126 (2003)
Jackson, D. A. & Pombo, A. Replicon clusters are stable units of chromosome structure: evidence that nuclear organization contributes to the efficient activation and propagation of S phase in human cells. J. Cell Biol. 140, 1285–1295 (1998)
The Cancer Genome Atlas Research Network. Integrated genomic characterization of endometrial carcinoma. Nature 497, 67–73 (2013)
Morrison, C. et al. The controlling role of ATM in homologous recombinational repair of DNA damage. EMBO J. 19, 463–471 (2000)
Parmar, K. et al. Hematopoietic stem cell defects in mice with deficiency of Fancd2 or Usp1. Stem Cells 28, 1186–1195 (2010)
Hu, Y. et al. PARP1-driven poly-ADP-ribosylation regulates BRCA1 function in homologous recombination mediated DNA repair. Cancer Discov. 4, 1430–1447 (2014)
Yang, D. et al. Association of BRCA1 and BRCA2 mutations with survival, chemotherapy sensitivity, and gene mutator phenotype in patients with ovarian cancer. J. Am. Med. Assoc. 306, 1557–1565 (2011)
Seki, M. et al. High-efficiency bypass of DNA damage by human DNA polymerase Q. EMBO J. 23, 4484–4494 (2004)
Seki, M. & Wood, R. D. DNA polymerase theta (POLQ) can extend from mismatches and from bases opposite a (6–4) photoproduct. DNA Repair 7, 119–127 (2008)
Zhang, Y. & Jasin, M. An essential role for CtIP in chromosomal translocation formation through an alternative end-joining pathway. Nature Struct. Mol. Biol. 18, 80–84 (2011)
Chiruvella, K. K., Liang, Z. & Wilson, T. E. Repair of double-strand breaks by end joining. Cold Spring Harb. Perspect. Biol. 5, a012757 (2013)
Alexandrov, L. B. et al. Signatures of mutational processes in human cancer. Nature 500, 415–421 (2013)
Lemée, F. et al. DNA polymerase theta up-regulation is associated with poor survival in breast cancer, perturbs DNA replication, and promotes genetic instability. Proc. Natl Acad. Sci. USA 107, 13390–13395 (2010)
Kang, J., D’Andrea, A. D. & Kozono, D. A DNA repair pathway-focused score for prediction of outcomes in ovarian cancer treated with platinum-based chemotherapy. J. Natl. Cancer Inst. 104, 670–681 (2012)
The Cancer Genome Atlas Network Comprehensive molecular portraits of human breast tumours. Nature 490, 61–70 (2012)
Ceccaldi, R. et al. Bone marrow failure in Fanconi anemia is triggered by an exacerbated p53/p21 DNA damage response that impairs hematopoietic stem and progenitor cells. Cell Stem Cell 11, 36–49 (2012)
Bennardo, N., Cheng, A., Huang, N. & Stark, J. M. Alternative-NHEJ is a mechanistically distinct pathway of mammalian chromosome break repair. PLoS Genet. 4, e1000110 (2008)
Yusufzai, T. & Kadonaga, J. T. HARP is an ATP-driven annealing helicase. Science 322, 748–750 (2008)
Barber, L. J. et al. RTEL1 maintains genomic stability by suppressing homologous recombination. Cell 135, 261–271 (2008)
Amunugama, R., Groden, J. & Fishel, R. The HsRAD51B–HsRAD51C stabilizes the HsRAD51 nucleoprotein filament. DNA Repair 12, 723–732 (2013)
Pitroda, S. P. et al. DNA repair pathway gene expression score correlates with repair proficiency and tumor sensitivity to chemotherapy. Sci. Transl. Med. 6, 229ra242 (2014)
Aggarwal, M., Sommers, J. A., Shoemaker, R. H. & Brosh, R. M., Jr Inhibition of helicase activity by a small molecule impairs Werner syndrome helicase (WRN) function in the cellular response to DNA damage or replication stress. Proc. Natl Acad. Sci. USA 108, 1525–1530 (2011)
Acknowledgements
We thank N. Shima, D. Chowdhury, G. Shapiro, J. Walter, D. Kozono, J. Ablain, M. Delord, J.-B. Lazaro and members of the D’Andrea laboratory for discussions. We also thank H. Kim for providing DNA constructs, L. Moldovan for providing GST tagged RAD51, S. Johnson for providing MDA-MB 436 cells, A. Y. Li for technical assistance, L. Moreau for chromosomal breakage analysis, and K. Mouw for critical reading of the manuscript. We thank H. Zhang for providing access to shRNA screening data shown in Extended Data Fig. 6i. We thank C. Cazaux and J.-S. Hoffmann for providing pcDNA Flag-tagged Polθ. R.C. received support from the Philippe Foundation and is a recipient of the Ovarian Cancer Research Fellowship (OCRF). This work was supported by NIH grants P50CA168504 and R01HL52725 and by grants from OCRF and BCRF.
Author information
Authors and Affiliations
Contributions
R.C. conceived the study, performed experiments, and wrote the manuscript. J.C.L. and T.Y. purified Polθ fragments from insect cells and performed ATPase and gel shift assays. R.A. performed D-loop formation assays. I.H. and S.J.E. performed the DNA fibres assay. B.P. performed mice work and analysed in vivo data. M.I.R.P. and S.J.B. performed the Polθ peptide array and the RAD51–ssDNA filament assembly and release assays. K.W.O. scored RAD51 foci. P.A.K. curated TCGA datasets for Figure 3a and Extended Data Figures 5h and 9e and provided clinical perspectives. A.D.D. conceived the study and wrote the manuscript. All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 POLQ is highly expressed in epithelial ovarian cancers (EOCs) and POLQ expression correlates with expression of HR genes.
a, b, Gene set enrichment analysis (GSEA) for expression of TransLesion Synthesis (TLS) (a) and polymerase (b) genes between primary cancers and control samples in 28 independent data sets from 19 different cancers types. Enrichment values (represented as a single dot for each gene in a defined data set) were determined using the rank metric score to compare expression values between cancers and control samples. Dots above the dashed line reflect enrichment in cancer samples, whereas dots below the dashed line show gene expression enriched in control samples. Data sets were ranked based on the amplitude of the rank metric score and plotted as shown. c, POLQ gene expression in 40 independent data sets from 19 different cancer types. For each data set, POLQ values were expressed as fold-change differences relative to the mean expression in control samples, which was arbitrarily set to 1. d, POLQ expression correlates with tumour grade and MKi67 gene expression in the ovarian TCGA (n = 494 patients with ovarian carcinoma (grade 1, n = 5; grade 2, n = 61; grade 3, n = 428) and control samples, n = 8). e, POLQ expression correlates with tumour grade MKi67 gene expression in the ovarian data set GSE9891 (n = 251 patients with ovarian serous and endometrious carcinoma for which grade status was available (grade 1, n = 20; grade 2, n = 88; grade 3, n = 143)). Statistical correlation was assessed using the Pearson test (for d: r = 0.65, P < 10−3; for e: r = 0.77, P < 10−3). f, Top-ranked biological pathways differentially expressed between samples expressing high levels of POLQ (high POLQ, first 33%, n = 95) relative to samples with low POLQ expression (low POLQ, 67%, n = 190) on the ovarian data set GSE9891 (n = 285 patients with ovarian carcinoma). Significance values were determined by the hypergeometrical test using the 200 most differentially expressed probe sets between the 2 groups (high POLQ and low POLQ). g, GSEA for expression of DNA repair genes between primary cancers and control samples in 5 independent ovarian cancer data sets. A representative heat map showing differential gene expression between ovarian cancers and controls is shown from GSE14407. For each data set, DNA repair genes were ranked based on the metric score reflecting their enrichment in cancer samples. The top 20 DNA repair genes primarily expressed in cancer samples compared to control samples is shown on the right. h, GSEA for the top 20 DNA repair genes defined in g between primary cancers and control samples in 40 independent cancer data sets. The nominal P value was used as a measure of the expression enrichment in cancer samples and represented as a waterfall plot. When the gene set expression was enriched in control samples, the P value was arbitrarily set to 1. i, POLQ expression correlates with RAD51 and FANCD2 gene expression in 285 samples from the ovarian data set GSE9891. Statistical correlation was assessed using the Pearson test (r = 0.71, P < 10−3). j, Top 10 genes that most closely correlated with POLQ expression (gene neighbours analysis) for 1,046 cell lines from the CCLE collection. DNA repair activity for these genes is indicated in the table. Increased HR gene expression is known to positively correlate with improved response to platinum based chemotherapy (a surrogate of HR deficiency) and thus can be predictive of decreased HR activity31,38. Conceptually, a state of HR deficiency may lead to compensatory increased expression of other HR genes. k, Top-ranked Gene Ontology (GO) terms for the molecular functions encoded by the top 20 DNA repair genes defined in Extended Data Fig. 1g. l, Schematic representation of Polθ domain structure with the helicases (BLM, RECQL4, RAD54B and RAD54L) that co-expressed with Polθ (from Extended Data Fig. 1g). Conserved amino-acid sequences of ATP binding and hydrolysis motifs (namely Walker A and B) are indicated. Cox plots in c show twenty-fifth to seventy-fifth percentiles, with lines indicating the median, and whiskers indicating the smallest and largest values. For d and e (top panels), each dot represents the expression value from one patient, brackets show mean ± s.e.m.
Extended Data Figure 2 Polθ is a RAD51-interacting protein required for maintenance of genomic stability.
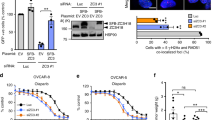
a, siRNA sequences (siPolθ1 and siPolθ2) efficiently downregulate exogenously transfected Polθ protein. Polθ levels were detected by immunoblotting with Flag or Polθ antibody (left) and by RT–qPCR using 2 different sets of POLQ primers (right). The asterisk on the immunoblot indicates a non-specific band. Expression was normalized using GAPDH as a reference gene. POLQ gene expression values are displayed as fold-change differences relative to the mean expression in control cells, which was arbitrarily set to 1. b, Quantification of baseline and HU-induced RAD51 foci in U2OS cells transfected with the indicated siRNA. c, Quantification of baseline and HU-induced γH2AX foci in U2OS cells transfected with the indicated siRNA. d, Quantification of IR-induced RAD51 foci in BrdU-positive U2OS cells transfected with the indicated siRNA. e, Polθ inhibition by siRNA induced a decrease in the cellular survival of 293T cells treated with MMC in a 3-day survival assay. f, Quantification of chromosomal aberrations in 293T cells transfected with the indicated siRNA. g, Schematic representation of Polθ truncation proteins used for RAD51 interaction studies. h, Endogenous RAD51 co-precipitates with Flag-tagged Polθ-ΔPol1 (Polθ-1–1,416) but not Polθ-1633-Cter, each stably expressed in HeLa cells. i, Sequence alignment between the RAD51-interacting motifs of C. elegans RFS-1 and human Polθ. j, Schematic of Polθ domain structure with its homologues HELQ and POLN. All data show mean ± s.e.m.
Extended Data Figure 3 Characterization of RAD51-interacting motifs in Polθ.
a, Substitution peptide array probed with recombinant RAD51 and analysed by immunoblotting. A 20-mer peptide spanning each of the RAD51 binding sites (shown in Fig. 1g) were created in which each amino acid of the original peptide was mutated to each of the 20 amino acids and RAD51 binding activity was tested. The amino acid change for each of the amino acids of the RAD51 interacting domain of Polθ is shown on the right. Ponceau staining was used to visualize position of the peptides within the array. b, GST–RAD51 pull-down with in vitro translated Polθ proteins missing indicated amino acids. c, Schematic of Polθ mutants used in complementation studies. d, Quantification of IR-induced RAD51 foci in U2OS cells stably integrated with empty vector (EV) or Polθ-ΔPol1 cDNA that is refractory to siPolθ1. Cells were transfected with indicated siRNA and subsequently treated with IR. The number of cells with more than 10 RAD51 foci was calculated relative to control cells (si Scr). e, DR-GFP assay in U2OS cells stably integrated with empty vector (EV) or indicated Polθ cDNA constructs refractory to siPolθ1 and transfected with indicated siRNA. All data show mean ± s.e.m.
Extended Data Figure 4 Polθ is an ATPase that suppresses RAD51–ssDNA nucleofilament assembly and formation of RAD51-dependent D-loop structures.
a, Representative ΔPol2 wild-type radiometric ATPase assay. b, Gel mobility shift assays with ΔPol2 wild type and ssDNA. c, Coomassie-stained gel showing the purified ΔPol2-A-dead fragment. d, Representative ΔPol2-A-dead radiometric ATPase assay. e, Quantification of ΔPol2-A-dead ATPase activity. (ssDNA, single-stranded DNA; dsDNA, double-stranded DNA). f, Assembly/disruption of RAD51-ssDNA filaments in the presence of increasing amounts of ΔPol2 wild type. The order in which each component was added to the reaction is noted above. g, Schematics of the formation of RAD51-dependent D-loop structures. h Formation of RAD51-containing D-loop structures following the addition of increasing amounts of ΔPol2 wild type. i, Fraction of D-loop formed following the addition of increasing amounts of ΔPol2 wild type. j, Effect of siPolθ and the different Polθ cDNA constructs on HR read-out. NA, not applicable. Data in i shows mean ± s.e.m.
Extended Data Figure 5 Polθ functions under replicative stress and is induced by HR deficiency.
a, Polθ recruitment to the chromatin is enhanced by UV treatment. HeLa cells stably integrated with either Flag-tagged ΔPol1 or Polθ-1633-Cter (Extended Data Fig. 2g) were subjected to UV treatment. Cells were collected at indicated time points after UV treatment and IPs were performed on nuclear and chromatin fractions. b, HeLa cells stably integrated with ΔPol1 were treated with UV and collected at the indicated time points following UV exposure. Polθ and RAD51 co-precipitation is enhanced by UV treatment. c, Quantification of DNA fibre lengths isolated from wild-type or Polq−/− MEFs. d, Quantification of DNA fibre lengths isolated from wild-type or Polq−/− MEFs transfected with either EV or Polθ cDNA constructs. e, POLQ gene expression was analysed by RT–qPCR in HR-deficient ovarian cancer cell lines (PEO-1 and UWB1-289) compared with other ovarian cancer cell lines, HeLa (cervical cancer) cells and 293T (transformed human embryonic kidney) cells. Expression was normalized using GAPDH gene as a reference. POLQ expression values are displayed as fold-change relative to the mean expression in HR-proficient control cells, which was arbitrarily set to 1. f, POLQ gene expression analysis (RT–qPCR) in 293T cells transfected with siRNA targeting FANCD2, BRCA1 or BRCA2 (left panel) and in corrected PD20 cells (PD20 + FANCD2) relative to FANCD2-deficient cells (PD20) (right panel). Expression was normalized using GAPDH gene as a reference. POLQ expression values are presented as fold-change relative to the mean expression in control cells, which was arbitrarily set to 1. g, POLQ gene expression in 5 data sets of serous epithelial ovarian carcinoma (frequently associated with an HR deficiency) and 1 data set of clear cell ovarian carcinoma (subgroup not associated with HR alterations). For each data set, POLQ expression values are displayed as fold-change differences relative to the mean expression in control samples, which was arbitrarily set to 1. h, Progression-free survival (PFS) after first line platinum chemotherapy for patients with ovarian carcinoma (ovarian carcinoma TCGA). Statistical significance was assessed by the log-rank test (P < 10−2). i, Effect of Polθ expression levels and HR status on tumour senstivity to cisplatin or PARPi. NA, not applicable. Box plots in c, d, and g show twenty-fifth to seventy-fifth percentiles, with lines indicating the median, and whiskers indicating the smallest and largest values. Data in e and f show mean ± s.e.m.
Extended Data Figure 6 Polθ inhibition sensitizes HR-deficient tumours to cytotoxic drug exposure.
a–c, Clonogenic formation of A2780 cells expressing scrambled (Scr) shRNA or shRNAs against FANCD2 or BRCA2 with increasing amounts of MMC (a), UV (b) or IR (c). d–f, Clonogenic formation of A2780 cells expressing scrambled (Scr) or FANCD2 shRNA, together with shRNA targeting Polθ, in increasing concentrations of CDDP (d), MMC (e) or PARPi (f). g, Inhibition of Polθ reduces the survival of A2780 cells after 3 days of continuous exposure to the ATM inhibitor Ku55933. h, Immunoblot analyses in A2780 cells expressing FANCD2 shRNA together with siRNA targeting Polθ or Scr at 24 h after indicated MMC pulse treatment. i, FANCA-deficient fibroblasts (GM6418) were infected with a whole-genome shRNA library and treated with MMC for 7 days. The fold-change enrichment of each shRNA after MMC treatment was determined by sequencing relative to the infected cells before treatment. TP53 depletion is known to improve survival of FANCA−/− cells33. WRN depletion has recently been shown to be synthetically lethal with HR deficiency39. Each column represents the mean of at least 2 independent shRNAs. All data show mean ± s.e.m.
Extended Data Figure 7 HR and Polθ repair pathways are synthetical lethal in vivo.
a, Genotypes frequencies of offspring from interbred Fancd2+/–Polq+/– mice. Ψ, four Fancd2−/−Polq−/− offsprings were observed with several congenital malformations and premature death within 48 h of birth. b, Description of Fancd2–/–Polq–/– offspring generated in the study. The offspring presented congenital malformations (that is, eye defects) together with reduced size and body weight. The arrow indicates absence of the right eye. c, Genotypes frequencies of E13.5 to E15 embryos (13.5 to 15 days post coitum) from interbred Fancd2+/–Polq+/– mice. d, Description of congenital malformations and their measured frequencies observed in E13.5 to E15 Fancd2–/–Polq–/– embryos generated in the study. e, Clonogenic formation of wild-type, Fancd2−/−, Polq−/− and Fancd2–/–Polq–/– MEFs with increasing concentrations of PARPi. f, A2780 cells were transduced with indicated shRNAs and xenotransplanted into both flanks of athymic nude mice. The tumour volumes for individual mice were measured biweekly for 8 weeks. Each group represents n ≥ 5 tumours from n ≥ 5 mice. g, Ki67 and γH2AX quantification in tumours treated with either vehicle or PARPi. h, Representative Ki67 and γH2AX staining of A2780-shFANCD2 xenografts expressing sh Scr or sh Polθ in athymic nude mice, treated with either vehicle or PARPi. Scale bars, 100 μM. i, In vivo competition assay design. j, Tumour chimaerism post-xenotransplantation for indicated conditions. k, Representative flow cytometry analysis of tumours before xenotransplantation (post-FACS sorting) or after xenotransplantation (post-transplant, PARPi). The percentage of GFP–RFP positive cells is indicated. l, Tumour chimaerism post-xenotransplantation for indicated conditions. For data in j and l, each circle represents data from one tumour and each group represents n ≥ 7 tumours from n ≥ 6 mice. Brackets show mean ± s.e.m. Data in e–g show mean ± s.e.m. For f each group represents n ≥ 6 tumours from n ≥ 6 mice.
Extended Data Figure 8 Polθ is required for HR-deficient cell survival and limits the formation of RAD51 structures in HR-deficient cells.
a, Clonogenic formation of Fancd2−/−Polq−/− MEFs transfected with full-length POLQ cDNA constructs in the presence of increasing concentrations of PARPi. b, Chromosome breakage analysis of FANCD2-depleted cells that were first transfected with the indicated siRNA and full-length POLQ cDNA constructs refractory to siPolθ1 and then exposed to MMC. c, DR-GFP assay in U2OS cells transfected with indicated siRNA. d, Quantification of baseline and IR-induced RAD51 foci in U2OS cells transfected with indicated siRNA. e, RAD51 recruitment to chromatin is enhanced by UV treatment. VU 423 cells (BRCA2−/−) were collected at indicated time points after UV treatment and immunoblotting performed on the cytoplasmic, nuclear and chromatin fractions. f, RAD51 recruitment to chromatin in VU 423 cells (BRCA2−/−) transfected with indicated siRNA. Histone H3 was used as a control for chromatin fractionation. All data show mean ± s.e.m.
Extended Data Figure 9 Polθ participates in error-prone DNA repair.
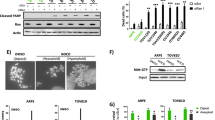
a, End-joining reporter assay in U2OS cells transfected with indicated siRNA and/or treated with PARPi. b, End-joining reporter assay in U2OS cells transfected with indicated siRNA and POLQ cDNA constructs refractory to siPolθ1. c, UV damage-induced Polθ foci formation in U2OS cells. Polθ foci were abolished by pre-treatment with PARPi. UT, untreated. d, Mutation frequency was determined in damaged supF plasmid, recovered from siRNA-treated 293T cells. e, Non-synonymous mutation count in ovarian, uterine and breast TCGA. All data show mean ± s.e.m.
Extended Data Figure 10 Model depicting the role of Polθ in DNA repair.
a, Mechanistic model for how Polθ limits RAD51–ssDNA filament assembly. According to this model, the ATPase domain of Polθ may prevent the assembly of RAD51 monomers into RAD51 polymers, perhaps by depleting local ATP concentrations. The RAD51 binding domains in the central region of Polθ may then sequester the RAD51 monomers, preventing filament assembly. b, (I) Under physiological conditions, Polθ expression is low and its impact on repair of DNA double-strand breaks (DSB) is limited. (II) When HR deficiency occurs, Polθ is then highly expressed and channels DSB repair towards alt-EJ. (III) In the case of an HR-defect, the loss of Polθ leads to cell death through the persistence of toxic RAD51 intermediates and inhibition of alt-EJ.
Supplementary information
Supplementary Table 1
This file contains the GEO accession number and details of the datasets used in the study. (XLSX 41 kb)
Supplementary Table 2
This file contains 200 probesets, differentially expressed between POLQ high and POLQ low expression groups. (XLSX 57 kb)
Supplementary Table 3
This file contains the Translesion Synthesis GeneSet and the Polymerase GeneSet. (XLSX 42 kb)
Rights and permissions
About this article

Cite this article
Ceccaldi, R., Liu, J., Amunugama, R. et al. Homologous-recombination-deficient tumours are dependent on Polθ-mediated repair. Nature 518, 258–262 (2015). https://doi.org/10.1038/nature14184
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14184
This article is cited by
-
POLQ inhibition attenuates the stemness and ferroptosis resistance in gastric cancer cells via downregulation of dihydroorotate dehydrogenase
Cell Death & Disease (2024)
-
The m6A demethylase FTO targets POLQ to promote ccRCC cell proliferation and genome stability maintenance
Journal of Cancer Research and Clinical Oncology (2024)
-
Genomic characterization of IDH-mutant astrocytoma progression to grade 4 in the treatment setting
Acta Neuropathologica Communications (2023)
-
CATI: an efficient gene integration method for rodent and primate embryos by MMEJ suppression
Genome Biology (2023)
-
Clinical application of PARP inhibitors in ovarian cancer: from molecular mechanisms to the current status
Journal of Ovarian Research (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
