
Abstract
Low-molecular-mass thiols in organisms are well known for their redox-relevant role in protection against various endogenous and exogenous stresses1,2,3. In eukaryotes and Gram-negative bacteria, the primary thiol is glutathione (GSH), a cysteinyl-containing tripeptide. In contrast, mycothiol (MSH), a cysteinyl pseudo-disaccharide, is dominant in Gram-positive actinobacteria, including antibiotic-producing actinomycetes and pathogenic mycobacteria. MSH is equivalent to GSH, either as a cofactor or as a substrate, in numerous biochemical processes4, most of which have not been characterized, largely due to the dearth of information concerning MSH-dependent proteins. Actinomycetes are able to produce another thiol, ergothioneine (EGT), a histidine betaine derivative that is widely assimilated by plants and animals for variable physiological activities5. The involvement of EGT in enzymatic reactions, however, lacks any precedent. Here we report that the unprecedented coupling of two bacterial thiols, MSH and EGT, has a constructive role in the biosynthesis of lincomycin A, a sulfur-containing lincosamide (C8 sugar) antibiotic that has been widely used for half a century to treat Gram-positive bacterial infections6,7,8,9. EGT acts as a carrier to template the molecular assembly, and MSH is the sulfur donor for lincomycin maturation after thiol exchange. These thiols function through two unusual S-glycosylations that program lincosamide transfer, activation and modification, providing the first paradigm for EGT-associated biochemical processes and for the poorly understood MSH-dependent biotransformations, a newly described model that is potentially common in the incorporation of sulfur, an element essential for life and ubiquitous in living systems.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Accession codes
References
Fahey, R. C. Glutathione analogs in prokaryotes. Biochim. Biophys. Acta 1830, 3182–3198 (2013)
Deponte, M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta 1830, 3217–3266 (2013)
Newton, G. L. et al. Bacillithiol is an antioxidant thiol produced in Bacilli. Nature Chem. Biol. 5, 625–627 (2009)
Rawat, M. & Av-Gay, Y. Mycothiol-dependent proteins in actinomycetes. FEMS Microbiol. Rev. 31, 278–292 (2007)
Cheah, I. K. & Halliwell, B. Ergothioneine; antioxidant potential, physiological function and role in disease. Biochim. Biophys. Acta 1822, 784–793 (2012)
Mason, D. J., Dietz, A. & DeBoer, C. Lincomycin, a new antibiotic. I. Discovery and biological properties. Antimicrob. Agents Chemother. 554–559 (1962)
Herr, R. R. & Bergy, M. E. Lincomycin, a new antibiotic. II. Isolation and characterization. Antimicrob. Agents Chemother. 560–554 (1962)
Lewis, C., Clapp, H. W. & Grady, J. E. In vitro and in vivo evaluation of lincomycin, a new antibiotic. Antimicrob. Agents Chemother. 570–582 (1963)
Mason, D. J. & Lewis, C. Biological activity of the lincomycin related antibiotics. Antimicrob. Agents Chemother. 7–12 (1964)
Newton, G. L., Buchmeier, N. & Fahey, R. C. Biosynthesis and functions of mycothiol, the unique protective thiol of Actinobacteria. Microbiol. Mol. Biol. Rev. 72, 471–494 (2008)
Jothivasan, V. K. & Hamilton, C. J. Mycothiol: synthesis, biosynthesis and biological functions of the major low molecular weight thiol in actinomycetes. Nature Prod. Rep. 25, 1091–1117 (2008)
Fan, F., Vetting, M. W., Frantom, P. A. & Blanchard, J. S. Structures and mechanisms of the mycothiol biosynthetic enzymes. Curr. Opin. Chem. Biol. 13, 451–459 (2009)
Reusser, F. Effect of lincomycin and clindamycin on peptide chain initiation. Antimicrob. Agents Chemother. 7, 32–37 (1975)
Wilson, D. N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nature Rev. Microbiol. 12, 35–48 (2014)
Peschke, U., Schmidt, H., Zhang, H.-Z. & Piepersberg, W. Molecular characterization of the lincomycin-production gene cluster of Streptomyces lincolnensis 78–11. Mol. Microbiol. 16, 1137–1156 (1995)
Koběrská, M. et al. Sequence analysis and heterologous expression of the lincomycin biosynthetic cluster of the type strain Streptomyces lincolnensis ATCC 25466. Folia Microbiol. 53, 395–401 (2008)
Scharf, D. H. et al. Epidithiol formation by an unprecedented twin carbon-sulfur lyase in the gliotoxin. Angew. Chem. Int. Ed. 124, 10211–10215 (2012)
Cooper, A. J. L. et al. Cysteine S-conjugate β-lyases: important roles in the metabolism of naturally occurring sulfur and selenium-containing compounds, exnobiotics, and anticancer agents. Amino Acids 41, 7–27 (2011)
Brahme, N. M. et al. Biosynthesis of the lincomycins. 2. Studies using stable isotopes on the biosynthesis of methylthiolincosaminide moiety of lincomycin A. J. Am. Chem. Soc. 106, 7878–7883 (1984)
Sasaki, E., Lin, C.-I., Lin, K.-Y. & Liu, H.-w. Construction of the octose 8-phosphate intermediate in lincomycin A biosynthesis: characterization of the reactions catalyzed by LmbR and LmbN. J. Am. Chem. Soc. 134, 17432–17435 (2012)
Lin, C.-I., Sasaki, E., Zhong, A. & Liu, H.-w. In vitro characterization of LmbK and LmbO: identification of GDP-D-erythro-α-D-gluco-octose as a key intermediate in lincomycin A biosynthesis. J. Am. Chem. Soc. 136, 906–909 (2014)
Newton, G. I., Leung, S. S., Wakabayashi, J. I., Rawat, M. & Fahey, R. C. The DinB superfamily includes novel mycothiol, bacillithiol, and glutathione S-transferases. Biochemistry 50, 10751–10760 (2011)
Feng, J. et al. The gene ncgl2918 encodes a novel maleylpyruvate isomerase that needs mycothiol as cofactor and links mycothiol biosynthesis and gentisate assimilation in Corynebacterium glutamicum. J. Biol. Chem. 281, 10778–10785 (2006)
Wang, R. et al. Crystal structures and site-directed mutagenesis of a mycothiol-dependent enzyme reveal a novel folding and molecular basis for mycothiol-mediated maleylpyruvate isomerization. J. Biol. Chem. 282, 16288–16294 (2007)
Seebeck, F. P. In vitro reconstitution of mycobacterial ergothioneine biosynthesis. J. Am. Chem. Soc. 132, 6632–6633 (2010)
Čermák, L. et al. Hybridization analysis and mapping of the celesticetin gene cluster revealed genes shared with lincomycin biosynthesis. Folia Microbiol. 52, 457–462 (2007)
Zhang, C. et al. Exploiting the reversibility of natural product glycosyltransferase- catalyzed reactions. Science 313, 1291–1294 (2006)
Kadlčík, S. et al. Adaptation of an l-proline adenylation domain to use 4-propyl-l-proline in the evolution of lincosamide biosynthesis. PLoS ONE 8, e84902 (2013)
Gantt, R. W., Peltier-Pain, P. & Thorson, J. S. Enzymatic methods for glyco (diversification/randomization) of drugs and small molecules. Natural Prod. Rep. 28, 1811–1853 (2011)
Sasaki, E. et al. Co-opting sulphur-carrier proteins from primary metabolic pathways for 2-thiosugar biosynthesis. Nature 510, 427–431 (2014)
Kieser, T. et al. Practical Streptomyces Genetics (John Innes Foundation, 2000)
Green, M. R. & Sambrook, J. Molecular Cloning: A Laboratory Manual 4th edn (Cold Spring Harbor Laboratory Press, 2012)
Fahey, R. C. & Newton, G. L. Determination of low molecular weight thiols using monobromobimane fluorescent labeling and high-performance liquid chromatography. Methods Enzymol. 143, 85–96 (1987)
Zhang, C. et al. Exploiting the reversibility of natural product glycosyltransferase- catalyzed reactions. Science 313, 1291–1294 (2006)
Lee, S. & Rossaza, J. P. First total synthesis of mycothiol and mycothiol disulfide. Org. Lett. 6, 365–368 (2004)
Ajayi, K., Thakur, V. V., Lapo, R. C. & Knapp, S. Intramolecular α-glucosaminidation: synthesis of mycothiol. Org. Lett. 12, 2630–2633 (2010)
Acknowledgements
This work was supported in part by grants from NSFC (81302674, 31430005, 91213303, 21472231 and 91413101), STCSM (14JC1407700 and 13XD1404500) and MST (2012AA02A706) of China.
Author information
Authors and Affiliations
Contributions
Q.Zhao, M.W. and Q.Zhang performed in vivo investigations. M.W. and D.X. prepared and characterized the chemical compounds. Q.Zhao and W.L. conducted sequence analysis. Q.Zhao and M.W. performed in vitro enzymatic investigations. Q.Zhao, M.W. and W.L. analysed the data and wrote the manuscript. W.L. directed the research.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
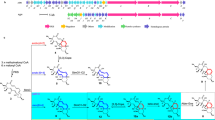
Extended Data Figure 2 Genes and/or clusters relevant to lincomycin biosynthesis in S. lincolnensis.
a, Biosynthetic gene cluster of lincomycin A. The mca homologue lmbE and two GTase-encoding genes, lmbV and lmbT, are shown in yellow, blue and green, respectively. The genes responsible for PPL incorporation (lmbC, lmbD and lmbN) are shown in grey. b, The location of mshAlin (shown in purple), which is not clustered with the other genes responsible for MSH biosynthesis, in the genome of S. lincolnensis. The flanking genes lin5590, lin5591, lin5593 and lin5594 share sequence homology with genes encoding the methylase (WP_026151264.1) from S. prunicolor, the DUF899-like protein (WP_004004056.1) from S. viridochromogenes, the YbjN-like protein (WP_020130332.1) from Streptomyces sp. 303MFCol5.2 and the hypothetical protein (WP_004000160.1) from S. viridochromogenes, respectively. c, The biosynthetic gene cluster of EGT. The gene egtDlin is shown in red.
Extended Data Figure 3 The production of MSH S-conjugate 1 and lincomycin A in various S. lincolnensis strains.
The strains include the mutants (i, for ΔlmbE; ii, for ΔlmbE-E80; iii, for ΔlmbE-E447; and iv, for ΔlmbE-E3457) and the wild-type control (v). For HPLC-MS analysis, the ESI m/z [M + H]+ modes are indicated in the dashed rectangle.
Extended Data Figure 4 Analysis of MSH and EGT production in the ΔmshAlin and ΔegtDlin mutant S. lincolnensis strains.
a, Derivatization of MSH and EGT with mBBr to generate the corresponding S-conjugates. b, HPLC-HR-MS analysis of MSH-mBBr (calculated for C27H41N4O14S+ [M + H]+ 677.2340) and EGT-mBBr (calculated for C19H26N5O4S+ [M + H]+ 420.1706) in the wild-type control (i), LL1005 (ii, ΔmshAlin mutant) and LL1010 (iii, ΔegtDlin mutant). c, The HR-ESI-MS spectra of EGT-mBBr (top) and MSH-mBBr (below).
Extended Data Figure 5 Phylogenetic analysis in the DinB-2 superfamily.
a, LmbV (from S. lincolnensis) and CcbV (from S. caelestis), shown in red, with selected DinB-2-like proteins (from various actinomycetes, in which the thiol MSH is dominant) in the phylogenetic tree. The evolutionary distances were computed using the p-distance method. The support for grouping clades i, ii, iii, iv and v (shaded in different colours) is indicated by bootstrap values. The known MSH-maleylpyruvate isomerase Ncgl2918 is shown in blue. b, Typical domain organization of the DinB-2-like proteins. The conserved DinB-2 domain is shown in green. Clade i features the C-terminal MDMPI-C domain. Proteins containing this domain include the MSH-maleylpyruvate isomerases, such as Ncgl2918. Clade ii features the C-terminal FGE-sulfatase domain, which is found in eukaryotic proteins required for post-translational modification to produce sulfatases, which are essential for the degradation and remodelling of sulfate esters. Clade iii features the N-terminal zf-HC2 domain, which contains a putative zinc-finger binding motif and is found in some anti-sigma factor proteins. Clade iv features the C-terminal SCP2 domain involved in binding sterols. Clade v features the C-terminal wyosine_f domain. Some proteins containing this domain appear to be important in wyosine base formation in a subset of phenylalanine-specific tRNAs. ‘Others’ indicate a number of DinB-2-like proteins that possess an unknown domain(s) either at the C terminus or at both the C and N termini of the proteins.
Extended Data Figure 6 Kinetic analysis of CcbV-catalysed thiol exchange.
a, pH dependence. The activity of CcbV in 50 mM PIPES (pH 6.0-7.0) or 50 mM Tris-HCl (pH 7.5–9.0) buffer was measured. b, c, Determination of the steady-state kinetic parameters for substrate 4 and for MSH, respectively. The error bars are standard error of mean (n = 3).
Extended Data Figure 7 Characterization of LmbT-catalysed reverse and forward glycosylation.
For HPLC-MS analysis, the ESI m/z [M + H]+ modes are indicated in dashed rectangle. a, Examination of the acylated C8-sugar transfer in the presence of LmbT, which showed that LmbT was unable to utilize 4 as a substrate for reverse glycosylation to generate the predicted GDP-D-α-d-sugar 7 (left) in the absence (top right) and in the presence (lower right) of LmbT. b, Characterization of LmbT-catalysed forward glycosylation. LmbT used 3 as a substrate for glycosylation to generate the GDP-d-α-d-sugar 5 (left) in the absence (top right) and in the presence (lower right) of LmbT.
Extended Data Figure 8 Characterization of PPL incorporation in lincomycin A biosynthesis.
For HPLC-MS analysis, the ESI m/z [M + H]+ modes are indicated in the dashed rectangle. a, In vivo product profiles of S. lincolnensis strains, including the mutants (i, for ΔlmbC; ii, for ΔlmbN; iii, for ΔlmbD; and iv, for ΔlmbT) and the wild-type control (v). b, Process of the incorporation of PPL (with EGT S-conjugate 5) into intermediate 4. PCP (blue), peptidyl carrier protein; and I (grey), isomerase. c, HPLC-MS examination of LmbC-catalysed conversion of holo-LmbN-PCP (m/z [M + H]+ calculated 11,842.94, found 11,843.42) into PPL-acylated LmbN-PCP (m/z [M + H]+ calculated 11,982.01, found 11,983.21) in the absence (left) and in the presence (right) of ATP. d, In vitro analysis of the condensation between PPL and 5 to generate 4. The catalyst systems included LmbC + CcbD (i), LmbC + LmbN (ii), LmbC + LmbN-PCP (iii), LmbC + LmbN + CcbD (iv), and LmbC + LmbN-PCP + CcbD (v).
Extended Data Figure 9 Kinetic analysis of LmbT-catalysed reversible glycosylation.
a, pH dependence. The activity of LmbT in 50 mM PIPES (pH 6.0–7.0) or 50 mM Tris-HCl (pH 7.5–9.0) buffer was measured. b, c, Determination of the steady-state kinetic parameters for substrate 5 and for GDP, respectively, in LmbT-catalysed reverse glycosylation. The error bars are standard error of mean (n = 3). d, Determination of the equilibrium constant (Keq) of LmbT-catalysed glycosylation. Keq = ([GDP]/[3]) × ([5]/[EGT]) = 1.94 × 1 = 1.94.
Extended Data Figure 10 Validation of the highly ordered process involving EGT-mediated assembly and MSH-associated post-modifications in lincomycin biosynthesis.
For HPLC-MS analysis, the ESI m/z [M + H]+ modes are indicated in the dashed rectangle. a, Examination of the thiol exchange using 5 as a substrate in the absence (top right) and in the presence (bottom right) of CcbV, showing that CcbV was able to convert 5 to MSH S-conjugate 6 (left). b, Determination of the hydrolysis reaction in the absence (top right) and in the presence (bottom right) of LmbE, showing that this enzyme was unable to utilize 6 as a substrate and convert it to the predicted mercapturic acid 8 (left).
Supplementary information
Supplementary Information
This file contains Supplementary Methods, Supplementary Text, Supplementary Figures 1-11, Supplementary Tables 1-7 and Supplementary References. (PDF 6271 kb)
Supplementary Data
This file contains Supplementary Table 8. (PDF 222 kb)
Supplementary Data
This file contains Supplementary Table 9. (PDF 287 kb)
Supplementary Data
This file contains Supplementary Table 10. (PDF 270 kb)
Rights and permissions
About this article

Cite this article
Zhao, Q., Wang, M., Xu, D. et al. Metabolic coupling of two small-molecule thiols programs the biosynthesis of lincomycin A. Nature 518, 115–119 (2015). https://doi.org/10.1038/nature14137
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14137
This article is cited by
-
Decreased Lincomycin B Content by Regulating Osmotic Pressure in Fermentation of Streptomyces lincolnensis
Biotechnology and Bioprocess Engineering (2023)
-
Thiocysteine lyases as polyketide synthase domains installing hydropersulfide into natural products and a hydropersulfide methyltransferase
Nature Communications (2021)
-
Comparative transcriptomic analysis reveals the significant pleiotropic regulatory effects of LmbU on lincomycin biosynthesis
Microbial Cell Factories (2020)
-
Enhanced lincomycin A production by calcium gluconate feeding in fermentation of Streptomyces lincolnensis
Bioresources and Bioprocessing (2019)
-
A diversity-oriented rhodamine library for wide-spectrum bactericidal agents with low inducible resistance against resistant pathogens
Nature Communications (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
