
Abstract
The early evolutionary history of Ornithodira (avian-line archosaurs) has hitherto been documented by incomplete (Lagerpeton1) or unusually specialized forms (pterosaurs and Silesaurus2). Recently, a variety of Silesaurus-like taxa have been reported from the Triassic period of both Gondwana and Laurasia, but their relationships to each other and to dinosaurs remain a subject of debate3,4,5. Here we report on a new avian-line archosaur from the early Middle Triassic (Anisian) of Tanzania. Phylogenetic analysis places Asilisaurus kongwe gen. et sp. nov. as an avian-line archosaur and a member of the Silesauridae, which is here considered the sister taxon to Dinosauria. Silesaurids were diverse and had a wide distribution by the Late Triassic, with a novel ornithodiran bauplan including leaf-shaped teeth, a beak-like lower jaw, long, gracile limbs, and a quadrupedal stance. Our analysis suggests that the dentition and diet of silesaurids, ornithischians and sauropodomorphs evolved independently from a plesiomorphic carnivorous form. As the oldest avian-line archosaur, Asilisaurus demonstrates the antiquity of both Ornithodira and the dinosaurian lineage. The initial diversification of Archosauria, previously documented by crocodilian-line archosaurs in the Anisian6, can now be shown to include a contemporaneous avian-line radiation. The unparalleled taxonomic diversity of the Manda archosaur assemblage indicates that archosaur diversification was well underway by the Middle Triassic or earlier.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

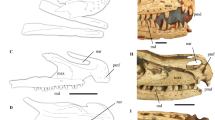
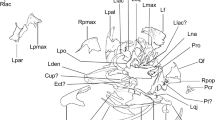
References
Romer, A. S. The Chañares (Argentina) Triassic reptile fauna. X. Two new but incompletely known long-limbed pseudosuchians. Breviora 378, 1–10 (1971)
Dzik, J. A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. J. Vertebr. Paleontol. 23, 556–574 (2003)
Ezcurra, M. D. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan & Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28, 649–684 (2006)
Irmis, R. B. et al. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317, 358–361 (2007)
Dzik, J. & Sulej, T. A review of the early Late Triassic Krasiejów biota from Silesia, Poland. Palaeontologia Polonica 64, 1–27 (2007)
Nesbitt, S. J. Arizonasaurus and its implications for archosaur divergence. Proc. R. Soc. Lond. B 270 (Suppl.) S234–S237 (2003)
Brusatte, S. L. et al. Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs. Science 321, 1485–1488 (2008)
Novas, F. E. Dinosaur monophyly. J. Vertebr. Paleontol. 16, 723–741 (1996)
Nesbitt, S. J. et al. Hindlimb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. J. Vertebr. Paleontol. 29, 498–516 (2009)
Wopfner, H. Tectonic and climatic events controlling deposition in Tanzanian Karoo basins. J. Afr. Earth Sci. 34, 167–177 (2002)
Haughton, S. H. On a collection of Karroo vertebrates from Tanganyika Territory. Q. J. Geol. Soc. Lond. 88, 634–671 (1932)
von Huene, F. Stenaulorhynchus, ein Rhynchosauride der ostafrikanischen Obertrias. Nova Acta Leopold. 6, 7–121 (1938)
von Huene, F. Ein kleiner Pseudosuchier und ein Saurischier aus den ostafrikanischen Mandaschichten. Neues Jb. Miner. Geol. Paläont. B 81, 61–69 (1939)
Crompton, A. W. On some Triassic cynodonts from Tanganyika. Proc. Zool. Soc. Lond. 125, 617–669 (1954)
Sidor, C., Angielczyk, K., Hopson, J., Nesbitt, S. & Smith, R. New vertebrate fossils from the Permo-Triassic Ruhuhu Basin of Tanzania. J. Vertebr. Paleontol. 28 (Suppl.). 142A–143A (2008)
Gay, S. A. & Cruickshank, A. R. I. Biostratigraphy of the Permian tetrapod faunas from the Ruhuhu Valley, Tanzania. J. Afr. Earth Sci. 29, 195–210 (1999)
Catuneanu, O. et al. The Karoo basins of south-central Africa. J. Afr. Earth Sci. 43, 211–253 (2005)
Rubidge, B. S. Re-uniting lost continents—fossil reptiles from the ancient Karoo and their wanderlust. S. Afr. J. Geol. 108, 135–172 (2005)
Ferigolo, J. & Langer, M. C. A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone. Hist. Biol. 19, 23–33 (2007)
Romer, A. S. The Chañares (Argentina) Triassic reptile fauna. XIV. Lewisuchus admixtus, gen. et sp. nov., a further thecodont from the Chañares beds. Breviora 390, 1–13 (1972)
Langer, M. C. & Benton, M. J. Early dinosaurs: a phylogenetic study. J. Syst. Palaeontology 4, 309–358 (2006)
Arcucci, A. Un nuevo Lagosuchidae (Thecodontia-Pseudosuchia) de la fauna de Los Chañares (edad reptil Chañarense, Triasico Medio), La Rioja, Argentina. Ameghiniana 24, 89–94 (1987)
Novas, F. E. The tibia and tarsus in Herrerasauridae (Dinosauria, incertae sedis) and the origin and evolution of the dinosaurian tarsus. J. Paleontol. 63, 677–690 (1989)
Nesbitt, S. J., Irmis, R. B. & Parker, W. G. A critical re-evaluation of the Late Triassic dinosaur taxa of North America. J. Syst. Palaeontology 5, 209–243 (2007)
Barrett, P. M. Prosauropod dinosaurs and iguanas: speculations on the diets of extinct reptiles. In Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record (ed. Sues, H.-D.) 42–78 (Cambridge Univ. Press, 2000)
Zanno, L. E., Gillette, D. D., Albright, L. B. & Titus, A. L. A new North American therizinosaurid and the role of herbivory in ‘predatory’ dinosaur evolution. Proc. R. Soc. B 276, 3505–3511 (2009)
Martinez, R. N. & Alcober, O. A. A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS One 4, e4397 (2009)
Charig, A. J. New Triassic archosaurs from Tanganyika including Mandasuchus and Teleocrater. Abstracts of Dissertations of the Univ. Cambridge 1955–56, 28–29 (1957)
Walker, A. D. A revision of the Jurassic crocodile Hallopus, with remarks on the classification of crocodiles. Phil. Trans. R. Soc. Lond. B 257, 323–372 (1970)
Butler, R. J. et al. A possible ctenosauriscid archosaur from the Middle Triassic Manda Beds of Tanzania. J. Vertebr. Paleontol. 29, 1022–1031 (2009)
Acknowledgements
We thank L. Herzog for preparation of the material and P. Barrett, P. Makovicky, A. Turner and N. Smith for discussions. We acknowledge A. Tibaijuka and C. Saanane for their help with permits and fieldwork logistics. Funding was provided by a National Geographic Society Research and Exploration grant (7587-05 to C.A.S.), the Evolving Earth Foundation (to S.J.N.), The Grainger Foundation (to K.D.A.), an NSF Postdoctoral Research Fellowship (DBI-0306158 to K.D.A.), and NSF Graduate Research Fellowships (to S.J.N. and R.B.I.).
Author Contributions S.J.N. and R.B.I. designed the research project; C.A.S. and K.D.A. designed the field project; S.J.N., C.A.S., K.D.A., R.M.H.S. and L.A.T. conducted fieldwork; S.J.N. described the material; R.M.H.S. interpreted the sedimentology and geology; S.J.N. and R.B.I. conducted the phylogenetic analysis; K.D.A. and R.B.I. conducted the comparative phylogenetic analyses; and S.J.N., R.B.I., C.A.S., R.M.H.S., L.A.T. and K.D.A. wrote the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
This file contains Supplementary Information: 1) Differential diagnosis for Asilisaurus kongwe, 2) Paratype and referred material of Asilisaurus kongwe, 3) Synonymy of Lewisuchus admixtus and Pseudolagosuchus major, 4) Details of the phylogenetic analysis, 5) Geology and taphonomy of the type locality, 6) Age of the Manda Beds, 7) Additional occurrences of the Silesauridae and 8) Details of Comparative Phylogenetic Tests. It also contains Supplementary Figures S1-S3 with Legends, Supplementary Tables S1- S2, and Supplementary References. (PDF 748 kb)
Rights and permissions
About this article
Cite this article
Nesbitt, S., Sidor, C., Irmis, R. et al. Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira. Nature 464, 95–98 (2010). https://doi.org/10.1038/nature08718
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/nature08718
This article is cited by
-
New reptile shows dinosaurs and pterosaurs evolved among diverse precursors
Nature (2023)
-
A new silesaurid from Carnian beds of Brazil fills a gap in the radiation of avian line archosaurs
Scientific Reports (2023)
-
The Late Triassic Ischigualasto Formation at Cerro Las Lajas (La Rioja, Argentina): fossil tetrapods, high-resolution chronostratigraphy, and faunal correlations
Scientific Reports (2020)
-
Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria
Nature (2020)
-
Dinosaur diversification linked with the Carnian Pluvial Episode
Nature Communications (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
