
Abstract
Arising from: A. Qvarnström, J. E. Brommer & L. Gustafsson Nature 441, 84–86 (2006); Qvarnström et al. reply
Qvarnström et al.1 test whether the preference of female collared flycatchers (Ficedula albicollis) for males with large forehead patches could have evolved as a by-product of selection acting on male patch size2. They find that the crucial genetic correlation between female choice and male patch size is not significant, and conclude that preference for large patches must have been shaped directly by selection. However, their use of the patch size of a female's social partner as a measure of choice is incomplete, and will result in low estimates of the potential for direct selection to shape female preference. Their study is therefore unable to resolve the question of how female preference for large forehead patches has evolved3.
Similar content being viewed by others
Main
Female preference for males with exaggerated ornaments is an important source of sexual selection4. For example, male collared flycatchers with enlarged forehead patches pair up with a female more rapidly than do normal males5. To resolve the controversy over whether such preferences can evolve as a by-product of selection acting on male ornaments3, Qvarnström et al.1 investigate the quantitative genetics underlying variation in female mate choice in relation to male forehead patch size. Whereas patch size is moderately heritable, the heritability of female choice is close to zero, with a non-significant genetic correlation between the two. They argue that their estimated genetic correlation from a free-living bird population provides a better test of the potential for indirect selection than the strong genetic correlations found from laboratory studies on fish6 and insects7. We challenge this view, and draw attention to important limitations in how Qvarnström et al. measured mate choice.
Whereas laboratory studies use the power of experimental choice trials, Qvarnström et al. use the patch size of a female's social partner. Believing that a female's social partner reflects her preference assumes that the distribution of male ornament size matches the distribution of female preference, and that every female can get what she wants. (This is akin to suggesting that Angelina Jolie is the only woman who finds Brad Pitt attractive and that most women prefer overweight men.) But in species such as the collared flycatcher, males and females are effectively removed from the pool of available partners when they form socially exclusive pair bonds. However, females mated to small-patched males do regularly copulate with extra-pair males with a larger patch, accounting for 90% of the sexual selection on male patch size8. By ignoring this important expression of preference, Qvarnström et al. overlook the main evolutionary pathway by which indirect selection on preference for large patches might act.
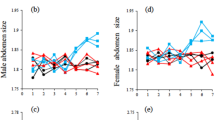
Given that mean female choice does not differ from mean male patch size and that variation in fitness is not larger in males than in females (Table 1 in ref. 1), the measure of choice used by Qvarnström et al. is unable to capture variation in female preference for large forehead patches. It therefore remains unclear whether variation in female preference affects which males females pair up with.
We agree with Qvarnström et al.1 that partnership formation is shaped by a range of ecological processes, if only because not every female can partner with her preferred male. However, this reduces the importance of female preference in partnership formation per se, and determines both her indirect and direct fitness. The potential for both indirect and direct selection to shape the evolution of preference is therefore, absolutely speaking, low. The issue about their relative importance cannot be settled until the potential for direct selection is quantified, and both are compared directly9. A non-significant genetic correlation between ornament and choice (between 10.3 and 0.3), and a significant heritability of choice (of 0.01–0.04) is not sufficient to support the argument.
Instead of dismissing results from the laboratory as unnatural, or from long-term wild data sets as unreplicated and correlational, we should study classic laboratory species in the wild and wild animals in captivity. This will allow for careful estimation of genetic variances and covariances of the appropriate traits, and thereby, for meaningful tests of evolutionary models of sexual selection10.
References
Qvarnström, A., Brommer, J. E. & Gustafsson, L. Nature 441, 84–86 (2006).
Kirkpatrick, M. & Ryan, M. J. Nature 350, 33–38 (1991).
Kokko, H., Brooks, R., Jennions, M. D. & Morley, J. Proc. R. Soc. Lond. B 270, 653–664 (2003).
Andersson, M. Sexual Selection (Princeton University Press, Princeton, New Jersey, 1994).
Qvarnström, A., Pärt, T. & Sheldon, B. C. Nature 405, 344–347 (2000).
Bakker, T. C. M. Nature 363, 255–257 (1993).
Iyengar, V. K., Reeve, H. K. & Eisner, T. Nature 419, 830–832 (2002).
Sheldon, B. C. & Ellegren, H. Anim. Behav. 57, 285–298 (1999).
Kirkpatrick, M. & Barton, N. H. Proc. Natl Acad. Sci. USA 94, 1282–1286 (1997).
Charmantier, A. & Sheldon, B. C. Trends Ecol. Evol. 21, 417–419 (2006).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Postma, E., Griffith, S. & Brooks, R. Evolution of mate choice in the wild. Nature 444, E16 (2006). https://doi.org/10.1038/nature05501
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature05501
This article is cited by
-
Evolution of mate choice in the wild (Reply)
Nature (2006)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
