
Abstract
Replying to Schliewen et al
We reported a case of sympatric speciation in the Nicaraguan Midas cichlid species complex1. Schliewen et al.2 question the interpretation of aspects of our data, but their proposed alternative scenario of multiple colonization and hybridization is considerably less parsimonious, contains some inconsistencies, and is incompatible with the available evidence.
Similar content being viewed by others

Main
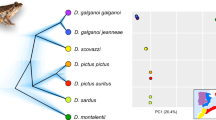
Amphilophus labiatus is not a sister species of the Lake Apoyo Amphilophus fauna3. The central haplotype in Fig. 2 of ref. 1 indeed contains specimens of A. labiatus and A. citrinellus; this figure, as indicated1, is a simplified version of our earlier one3. However, we have shown that A. labiatus is more distantly related to the monophyletic Lake Apoyo assemblage than is A. citrinellus from Lake Nicaragua3 (Nature did not permit us to show additional analyses or figures to this effect). This is also supported by morphometrics4 and the absence of A. labiatus from Lake Apoyo. Our microsatellite, mitochondrial (mt) DNA and amplified fragment-length polymorphism (AFLP) analyses1 confirm that A. zaliosus and A. citrinellus from Lake Apoyo are each other's closest relatives.
There is no evidence to support the assertion by Schliewen et al.2 that A. citrinellus of Lake Apoyo is closer to A. citrinellus of Lake Nicaragua than is A. zaliosus. Factorial correspondence analysis does not either, as it illustrates present but not past genetic distances (in fact, any ancestral population should be equidistant from all of its descendants). Similarly, with only three potential cases in the more than 120 individuals included from Lake Apoyo (as determined by the analyses using Structure software; see Fig 3 in ref. 1), introgression is very rare in A. citrinellus — if it exists at all (P < 80%), as determined by the Structure analysis. The argument by Schliewen et al.2 for secondary introgression from Lake Nicaragua into Lake Apoyo is based on a single specimen, which is unlikely to be an introgressant as it contains alleles of the genomes of all three populations, which is likely to be an artefact of the analysis. The monophyly of Lake Apoyo's Amphilophus species and the complete endemism of its mtDNA haplotypes argue against secondary colonization.
Instead, the analyses all indicate that A. zaliosus evolved sympatrically from A. citrinellus within Lake Apoyo. We showed that A. zaliosus is only about half as old as A. citrinellus from Lake Apoyo1 (note that A. citrinellus from Lake Apoyo carries only a subset of the global A. citrinellus microsatellite alleles and that A. zaliosus carries only about half of the Lake Apoyo A. citrinellus alleles). These genetic data therefore rule out the alternative scenario proposed by Schliewen et al., in which A. citrinellus entered Lake Apoyo in a second wave of colonization after A. zaliosus.
We do not believe that our sampling of the taxonomic diversity in Lake Apoyo was inadequate. Our Lake Apoyo data set does include morphs that others5 call “chancho”, “short” and “amarillo”. These morphotypes have never been formally described as species, no voucher specimens and no phenotypic or meristic information is available (only some photographs), and no experimental or observational data have been published that would support assortative mating. A previous genetic analysis5 based on three microsatellites yielded inconclusive results. Our own, much more detailed, analyses1 find, so far, evidence for only two genetically discernable units of Amphilophus in Lake Apoyo — A. zaliosus and A. citrinellus. The seeming overlap in morphospace between the two Lake Apoyo species is due only to the two-dimensional projection of a multidimensional plot.
In summary, we maintain that the data fully support our original interpretations, whereas Schliewen et al.2 propose a much less likely scenario that is not supported by the available data.
References
Barluenga, M., Stolting, K. N., Salzburger, W., Muschick, M. & Meyer, A. Nature 439, 719–723 (2006).
Schliewen, U. K., Kocher, T. D., McKaye, K. R., Seehausen, O. & Tautz, D. Nature 444, doi:10.1038/nature05419 (2006).
Barluenga, M. & Meyer, A. Mol. Ecol. 13, 2061–2076 (2004).
Klingenberg, C. P., Barluenga, M. & Meyer, A. Biol. J. Linn. Soc. 80, 397–408 (2003).
McKaye, K. R. et al. Cuadernos de Investigación de la UCA 12, 19–47 (2002).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Barluenga, M., Stölting, K., Salzburger, W. et al. Evidence for sympatric speciation? (Reply). Nature 444, E13 (2006). https://doi.org/10.1038/nature05420
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature05420
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
