
Abstract
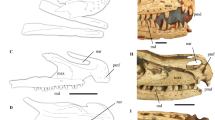
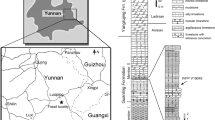
Discovering evidence of behaviour in fossilized vertebrates is rare. Even rarer is evidence of behaviour in non-avialan dinosaurs that directly relates to stereotypical behaviour seen in extant birds (avians) and not previously predicted in non-avialan dinosaurs1,2. Here we report the discovery of a new troodontid taxon from the Early Cretaceous Yixian Formation of western Liaoning, China. Numerous other three-dimensionally preserved vertebrate fossils have been recovered recently at this locality, including some specimens preserving behavioural information3. The new troodontid preserves several features that have been implicated in avialan origins. Notably, the specimen is preserved in the stereotypical sleeping or resting posture found in extant Aves4. Evidence of this behaviour outside of the crown group Aves further demonstrates that many bird features occurred early in dinosaurian evolution5,6.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others

References
Norell, M. A., Clark, J. M., Chiappe, L. M. & Dashzeveg, D. A nesting dinosaur. Nature 378, 774–776 (1995)
Varricchio, D. J., Jackson, F., Borkowski, J. J. & Horner, J. R. Nest and egg clutches of the dinosaur Troodon formosus and the evolution of avian reproductive traits. Nature 385, 247–250 (1997)
Meng, Q.-J. Liu, J. Y., Varricchio, D. J., Huang, T. & Gao, C.-L. Parential care in an ornithischian dinosaur. Nature 431, 145–146 (2004)
Wing, L. W. Natural History of Birds (Ronald Press, New York, 1956)
Xu, X. Deinonychosaurian fossils from the Jehol Group of western Liaoning and the coelurosaurian evolution. PhD dissertation, Chinese Academy of Sciences (2002)
Sereno, P. C. The evolution of dinosaurs. Science 284, 2137–2147 (1999)
Swisher, C. C. III et al. Further support for a Cretaceous age for the feathered-dinosaur beds of Liaoning, China: New 40Ar/39Ar dating of the Yixian and Tuchengzi Formations. Chin. Sci. Bull. 47, 135–138 (2002)
Xu, X., Zhou, Z. H. & Wang, X. L. The smallest known non-avian theropod dinosaur. Nature 408, 705–708 (2000)
Elzanowski, A. in Mesozoic Birds: Above the Heads of Dinosaurs (eds Chiappe, L. M. & Witmer, L. M.) 129–159 (Univ. California Press, Berkeley, 2002)
Jones, T. D., Farlow, J. O., Ruben, J. A., Henderson, D. M. & Hillenius, W. J. Cursoriality in bipedal archosaurs. Nature 406, 716–718 (2000)
Russell, D. A. & Dong, Z.-M. A nearly complete skeleton of a new troodontid dinosaur from the Early Cretaceous of the Ordos Basin, Inner Mongolia, People's Republic of China. Can. J. Earth Sci. 30, 2163–2173 (1994)
Currie, P. J. & Dong, Z. New information on Cretaceous troodontids (Dinosauria, Theropoda) from the People's Republic of China. Can. J. Earth Sci. 38, 1753–1766 (2001)
Currie, P. J. Bird-like characteristics of the jaws and teeth of troodontid theropods (Dinosauria: Saurischia). J. Vert. Paleontol. 7, 72–81 (1987)
Norell, M. A., Makovicky, P. J. & Clark, J. M. A new troodontid theropod from Ukhaa Tolgod, Mongolia. J. Vert. Paleontol. 20, 7–11 (2001)
Makovicky, P. J., Norell, M. A., Clark, J. M. & Rowe, T. Osteology and relationships of Byronosaurus jaffei (Theropda: Troodontidae). Am. Mus. Novit. 3402, 1–32 (2003)
Xu, X., Norell, M. A., Wang, X. L., Makovicky, P. J. & Wu, X. C. A basal troodontid from the Early Cretaceous of China. Nature 415, 780–784 (2002)
Hwang, S. H., Norell, M. A., Ji, Q. & Gao, K. New specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from northeastern China. Am. Mus. Novit. 3381, 1–44 (2002)
Norell, M. & Makovicky, P. J. Important features of the dromaeosaur skeleton: information from a new specimen. Am. Mus. Novit. 3215, 1–28 (1997)
Chiappe, L. M., Ji, S.-A., Ji, Q. & Norell, M. A. Anatomy and systematics of the Confuciusornithidae (Theropod: Aves) from the late Mesozoic of Northeastern China. Bull. Am. Mus. 242, 1–89 (1999)
Chiappe, L. M., Norell, M. A. & Clark, J. M. The skull of a relative of the stem-group bird Mononykus. Nature 392, 275–282 (1998)
Clark, J. M., Norell, M. A. & Chiappe, L. M. An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest. Am. Mus. Novit. 3265, 1–35 (1999)
Hwang, S. H., Norell, M. A., Ji, Q. & Gao, K.-Q. A large compsognathid from the Early Cretaceous Yixian Formation of China. J. Syst. Palaentol. 2, 13–30 (2004)
Barsbold, R. & Osmolska, H. The skull of Velociraptor (Theropoda) from the Late Cretaceous of Mongolia. Acta Palaeontol. Polonica 44, 189–219 (1999)
Hill, R. W., Beaver, D. L. & Veghte, J. H. Body surface temperatures and thermoregulation in the black-capped chickadee (Parus atricapillus). Physiol. Zool. 53, 305–321 (1980)
Marsh, R. L. & Dawson, W. R. Avian adjustments to cold. Adv. Comp. Environ. Physiol. 4, 205–253 (1989)
Chang, M.-M., Chen, P.-J., Wang, Y.-Q., Wang, Y. & Miao, D.-S. The Jehol Biota (Shanghai Scientific & Technical Publishers, Shanghai, 2003)
Guo, Z., Liu, J. & Wang, X. Effect of Mesozoic volcanic eruptions in the western Liaoning Province, China on paleoclimate and paleoenvironment. Sci. Chin. Ser. D 46, 1261–1272 (2003)
Loop, D. B., Dingus, L., Swisher, C. C. III & Minjin, C. Life and death in a Late Cretaceous dune field, Nemegt basin, Mongolia. Geology 26, 27–30 (1998)
Ji, Q. et al. An early ostrich dinosaur and implications for ornithomimosaur phylogeny. Am. Mus. Novit. 3420, 1–19 (2003)
Schweitzer, M. H. & Marshall, C. L. A molecular model for the evolution of endothermy in the theropod-bird lineage. J. Exp. Zool. 291, 317–338 (2001)
Acknowledgements
We thank J. Meng, X.-J. Ni, Y.-M. Hu, J. Clarke and R. Prum for discussions, J.-M. Yang and H.-J. Wang for preparing the specimen, and M. Ellison for illustrations. We also thank Z.-H. Zhou and other members of the Liaoxi expedition team of the IVPP. This work was supported by grants from the National Natural Science Foundation of China, Special Funds for Major State Basic Research Projects of China, National Geographic Society, American Museum of Natural History, Chinese Academy of Sciences and National Science Foundation of USA.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing financial interests.
Supplementary information
Supplementary Data
Files include: (1) character list; (2) data matrix; (3) analysis results (including a figure); (4) lengths of elements in Mei long; (5) relative proportions of elements in some avian and non-avian theropods. (DOC 406 kb)
Rights and permissions
About this article
Cite this article
Xu, X., Norell, M. A new troodontid dinosaur from China with avian-like sleeping posture. Nature 431, 838–841 (2004). https://doi.org/10.1038/nature02898
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/nature02898
This article is cited by
-
Conserved in-ovo cranial ossification sequences of extant saurians allow estimation of embryonic dinosaur developmental stages
Scientific Reports (2020)
-
Hummingbird-sized dinosaur from the Cretaceous period of Myanmar
Nature (2020)
-
Study on the Jehol Biota: Recent advances and future prospects
Science China Earth Sciences (2020)
-
Halszkaraptor escuilliei and the evolution of the paravian bauplan
Scientific Reports (2019)
-
A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution
Nature Communications (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
