
Abstract
Containment of highly lethal Ebola virus outbreaks poses a serious public health challenge. Although an experimental vaccine has successfully protected non-human primates against disease1, more than six months was required to complete the immunizations, making it impractical to limit an acute epidemic. Here, we report the development of accelerated vaccination against Ebola virus in non-human primates. The antibody response to immunization with an adenoviral (ADV) vector encoding the Ebola glycoprotein (GP) was induced more rapidly than with DNA priming and ADV boosting, but it was of lower magnitude. To determine whether this earlier immune response could nonetheless protect against disease, cynomolgus macaques were challenged with Ebola virus after vaccination with ADV–GP and nucleoprotein (NP) vectors. Protection was highly effective and correlated with the generation of Ebola-specific CD8+ T-cell and antibody responses. Even when animals were immunized once with ADV–GP/NP and challenged 28 days later, they remained resistant to challenge with either low or high doses of virus. This accelerated vaccine provides an intervention that may help to limit the epidemic spread of Ebola, and is applicable to other viruses.
Similar content being viewed by others

Main
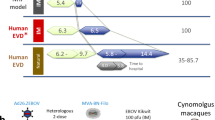
Mice were immunized with plasmid DNA encoding Ebola GP, the trimeric virion-associated glycoprotein2 involved in cellular pathogenicity3,4,5,6, followed by boosting with ADV–GP, or with ADV–GP only. The antibody response, a surrogate for protection1,7, was measured using an enzyme-linked immunosorbent assay (ELISA). After DNA vaccination, titres were modest but increased 100- to 1,000-fold with ADV–GP boosting (Fig. 1a). In contrast, vaccination with ADV–GP gave rise to a lower antibody titre, but it was generated more rapidly. To investigate whether immunization with adenoviral vectors alone might protect against Ebola virus infection, alternative immunization schedules in macaques were developed for comparison to the previous DNA/ADV protocol (Fig. 1b, middle and bottom panels compared with top panel).

a, The time course of Ebola-specific antibody responses by DNA prime and adenovirus boost compared with adenoviral immunization alone is shown (see Methods). Data represent the relative ELISA titre to Ebola GP after immunization with DNA/ADV–GP or ADV–GP/ADV–GP in BALB/c mice using a log scale. b, Immunization schedule for previously used heterologous prime–boost vaccine (top), adenoviral prime and boost (middle), and single adenoviral virus (bottom) immunizations. Challenge was performed with a 1995 isolate of Ebola virus (Zaire) at 32, 10 or 4 weeks after the initial immunization, respectively.
Cynomolgus macaques were immunized with ADV–GP and ADV–NP, followed by boosting 9 weeks later (Fig. 1b, middle panel). One week after the boost, animals were challenged with either a low (13 plaque-forming units (PFUs)) or high (1,500 PFUs) dose of a 1995 isolate of Ebola virus Zaire. These doses were uniformly fatal 6–12 days afterwards in saline-injected control animals. In contrast, the ADV–GP/NP immunized monkeys (n = 4) were completely protected, confirmed by viral load (Fig. 2). Analysis of the cell-mediated and humoral immune responses revealed significant increases in the CD8+ T-cell response to Ebola antigens by intracellular cytokine staining for interferon (IFN)-γ, seen before exposure to virus, in contrast to control animals where no response was seen (Fig. 3a). Similarly, antibody titres to the virus were stimulated in vaccinated animals, which minimally increased after the viral challenge (Fig. 3b). No substantial increases were observed in the numbers of Ebola-specific CD4+ T cells at this time (data not shown). Both CD8+ cellular and humoral immune responses therefore were associated with protection.

Plasma viraemia in monkeys after infection with Ebola virus. Asterisks represent the time of death in control animals. The data represent the reciprocal endpoint dilution of serum for each monkey. Results are shown for four immunized animals challenged with Ebola Zaire at 13 PFUs (low dose; filled symbols, left), four immunized animals challenged at 1,500 PFUs (high dose; filled symbols, right), and five saline-injected control animals (open symbols).

a, Intracellular flow cytometry was performed to quantify IFN-γ production from Ebola-specific CD8 lymphocytes from saline injected (control) or ADV–GP/NP immunized (subject) monkeys at weeks 0 and 9. Immune responses before (day 0) and after (days 3, 6) challenge at week 10 are shown for CD8 cells. No substantial increases were observed in the CD4 population. Non-stimulated cells gave responses similar to those of the control subjects, at background levels. The gating strategy for FACs analysis is similar to previous analyses (Supplementary Information). b, ELISA titres of Ebola-specific antibodies in serum of vaccinated animals collected at week 0 (pre-immune, left), week 9 (pre-boost, middle) and week 10 (day of challenge, right) relative to the time of the first immunization. ELISA results represent endpoint dilution titres determined by optical density as described in Methods.
A second adenoviral immunization did not substantially increase the Ebola-specific immune responses (data not shown), raising the notion that the primary immunization was sufficient to confer protection. To address this possibility, a single immunization was given, and animals were challenged one month afterwards (Fig. 1b, bottom panel). Both at low and high viral challenge doses, animals were completely protected against infection (Fig. 4a). In this case, changes in the intracellular IFN-γ response in T lymphocytes were not consistently seen (data not shown); however, Ebola-specific T-cell responses were detected with intracellular tumour-necrosis factor (TNF)-α. CD8 responses were observed before challenge or were induced soon thereafter in five of eight animals, once again correlating with protection against infection (Fig. 4b, right). In contrast, CD4+ responses, not detectable before inoculation, increased after challenge (Fig. 4b, left). Immunoglobulin-γ (IgG) antibody titres, readily detected at the time of inoculation, were also associated with protection (Fig. 4c). These data demonstrated that a single ADV–GP/NP injection can accelerate vaccine protection and long-term survival against Ebola in non-human primates (Fig. 4d).

a, Immunization and challenge were performed with the 1995 Zaire subtype Ebola virus as in Fig. 1b (bottom), and plasma viraemia in monkeys after challenge was measured as above (see Fig. 2 legend) for four immunized animals inoculated with 18 PFUs (low dose; filled symbols, left) and four animals injected with 1,762 PFUs (high dose; filled symbols, right) or two saline-injected controls (open symbols). b, Intracellular flow cytometry was performed using antibodies to TNF-α in CD4 and CD8 lymphocytes from immunized monkeys (subject), each panel representing an individual macaque. Immune responses before (day 0) and after (days 6, 10) challenge on day 28 are shown. Horizontal bars indicate the average value per group, and filled circles represent values for individual subjects. c, Endpoint dilution ELISA titres of Ebola-specific antibodies in serum collected two weeks after immunization with ADV–GP/NP, determined by optical density as described in Methods. d, Kaplan–Meier survival curve of macaques, immunized as indicated, and challenged with a low or high dose of PFUs of Ebola virus.
Ebola virus infection is characterized by its rapid onset, high person-to-person transmissibility, and significant mortality rate. The mainstay of treatment has been supportive therapy, and prevention has been dependent on containment using barrier precautions. Effective protection was achieved previously in primates with a heterologous DNA prime and adenoviral boost strategy. The prime–boost immunization relies on the ability of the adenoviral boost to expand the primary T-cell response induced by DNA vaccination. When animals are primed with ADV vectors alone, a robust Ebola-virus-specific cellular and humoral immune response is more rapidly achieved, although the response to a second ADV–GP/NP injection is blunted, probably because of anti-vector immunity. Here, we explored the possibility that this more rapid initial immune response may nonetheless confer protection and outweigh the stronger immune response that requires additional time. A single immunization with an adenoviral vector encoding Ebola virus proteins is sufficient to confer protection against lethal challenge within four weeks, and this response correlates with both cellular and humoral immune responses to the infection.
Although antibody titres correlated here with the protective response, previous studies in non-human primates have suggested that the passive transfer of antibody is insufficient to provide long-lasting protection against Ebola virus8. In rodent studies with adapted Ebola virus, passive transfer of antibodies9,10 or adoptive transfer of cytotoxic T cells11 showed protection when given before infection. A more sensitive but less quantitative CD4 lympho-proliferative response correlated with protection in the previous DNA/ADV prime–boost study, in which CD8 responses were not measured1. In addition to the antibody response induced by the vaccine in the present study, both CD4 and CD8 responses were observed after the challenge. The fact that CD4 responses were not observed before challenge in either protocol whereas CD8 responses were more consistently seen beforehand suggests that the CD8 response is likely to have an important role in protection in non-human primates, but further analysis will be required to assess the relative importance of the cellular and humoral immune responses in the mechanism of protection.
The approach to single vaccine injection with ADV vectors is relevant to the containment and treatment of Ebola virus and related outbreaks that are continuing to emerge in central Africa12. Should this vaccine approach prove effective in humans, our finding raises the possibility that ring vaccination could be used to contain outbreaks, similar to smallpox in the past. This result also suggests alternative strategies for vaccination against Ebola or other acute pathogenic diseases. The prime–boost strategy remains more immunologically potent and, if the response is highly durable, may still be useful for preventative vaccines, for example, in hospital workers. In contrast, the single adenoviral vaccine administration may be better used during acute outbreaks. It is also possible that alternative viral vectors, such as those derived from other adenovirus serotypes or from poxvirus vectors, might be used to boost an ADV type 5 vector primary immunization. Alternative ADV serotypes will also help to overcome immunity to natural ADV type 5 infection that could potentially reduce vaccine efficacy in some populations. A one-shot vaccine may be helpful in the control of Ebola virus outbreaks in great ape populations of central Africa13. Analogous single-dose ADV vector immunization could also be used for other emerging and highly lethal infectious pathogens, such as Marburg, Lassa or the SARS coronavirus.
Methods
Vector construction
ADV–GP and ADV–NP were prepared as described previously1. The recombinant adenoviral vector was made according to previously published methods14. A dose of 1010 (mice) or 1012 (non-human primates) adenoviral vector particles for each component was administered to each animal without adverse effects.
Animal study and safety
Twenty cynomolgus macaques (Macaca fascicularis), 3 yr old and weighing 2–3 kg, obtained from Covance, were used for immunization and challenge experiments. The monkeys, housed singly, were anaesthetized with ketamine to obtain blood specimens and to administer vaccines. They received regular enrichment according to the Guide for the Care and Use of Laboratory Animals (DHEW number NIH 86-23). Before Ebola virus challenge and to the end of each experiment, the animals were maintained in the Maximum Containment Laboratory (BSL-4) and fed and checked daily.
Mouse immunization
DNA and adenoviral vectors expressing Ebola Zaire glycoprotein (Mayinga strain) were constructed as described previously7,15 with gene expression under control of the cytomegalovirus enhancer and promoter in the plasmid. Mice (n = 10 per group) were immunized intramuscularly with 100 µg DNA (pGP) and/or 1010 particles of adenovirus (ADV–GP). DNA vaccination was performed on days 0, 14 and 24 with adenoviral boost on day 42. Adenoviral injection was performed on days 0 and 42, and samples were collected for ELISA titres at the indicated times. ELISA IgG titres were determined using 96-well plates as previously described16, and specific antigen binding was detected using a goat anti-human IgG (H + L)-horseradish conjugate and ABTS/peroxide (substrate/indicator).
ELISA
Polyvinyl chloride ELISA plates (Dynatech) were coated with 50 µl antigen per well and incubated overnight at 4 °C. All further incubations were carried out at room temperature. The antigen used was purified Ebola virus (about 1 mg ml-1 total protein) inactivated by gamma irradiation. Plates were then washed five times with PBS containing Tween-20. Test sera were diluted in half-log concentrations from 1:31.6 through to 1:100,000 and allowed to react with the antigen-coated wells for 60 min. After washing plates five times, goat anti-monkey IgG (whole molecule; ICN Biomedicals) conjugated to horseradish peroxidase was used as a detection antibody. Bound IgG was detected by 2,2′-azinobis-[3-ethylbenzothizoline-6-sulphonic acid] diammonium salt and the optical density was determined. A panel of normal serum was run each time the assay was performed. A cut-off value for a positive result was calculated as the mean optical density (at a 1:100 dilution) for the normal sera plus 3 standard deviations.
Intracellular cytokine analysis
Peripheral blood mononuclear cells were isolated from cynomolgus macaque whole-blood samples by separation over Ficoll. Approximately 1 × 106 cells were stimulated in 200 µl RPMI medium (GIBCO) for 6 h at 37 °C with anti-CD28 and anti-CD49d antibodies and either DMSO or a pool of 15-nucleotide peptides spanning the Ebola GP Zaire (Mayinga strain) open reading frame. The peptides were 15 nucleotides overlapping by 11 spanning the entire Ebola glycoprotein at a final concentration of 2 µg ml-1. Cells were fixed and permeablized with FACS lyse (Becton Dickinson) supplemented with Tween-20, and stained with a mixture of antibodies against lineage markers (CD3-PE, CD4-PerCP, CD8-FITC) and either TNF-APC or IFN-γ-APC. Samples were run on a FACS Calibur and analysed using the software FlowJo. Positive gating for lymphocytes using forward versus side scatter was followed by CD3+/CD8- and CD3+/CD4- gating, and specific populations were further defined by anti-CD4 and anti-CD8 positivity, respectively. Cytokine-positive cells were defined as a percentage within these individual lymphocyte subsets, and at least 200,000 events were analysed for each sample.
Macaque immunization
In conducting this research, the investigators adhered to the Guide for the Care and Use of Laboratory Animals, prepared by the Institute of Laboratory Animal Resources, National Research Council. The facilities are fully accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International. Cynomolgus macaques were injected intramuscularly at the indicated times (Fig. 1b) with an equal mixture of 2 × 1012 particles of ADV–GP and ADV–NP. Viral challenge was performed by inoculation of animals in the left or right caudal thigh with 0.5 ml of viral stock that contained a target dose of either about 10 or about 1,000 PFUs of Ebola virus (Zaire species) at ten weeks (Fig. 2) or four weeks (Fig. 4) after the initial immunization, and actual titre was confirmed by plaquing. No adverse effects of the adenovirus vaccination were observed acutely. The Ebola virus used in this study was originally obtained from a fatally infected human from the former Zaire in 1995 (ref. 17). Collection of serum and blood for viral load and ELISA titres was performed as previously described1.
References
Sullivan, N. J., Sanchez, A., Rollin, P. E., Yang, Z.-Y. & Nabel, G. J. Development of a preventive vaccine for Ebola virus infection in primates. Nature 408, 605–609 (2000)
Sanchez, A. et al. Biochemical analysis of the secreted and virion glycoproteins of Ebola virus. J. Virol. 72, 6442–6447 (1998)
Yang, Z.-Y. et al. Identification of the Ebola virus glycoprotein as the main viral determinant of vascular cell cytotoxicity and injury. Nature Med. 6, 886–889 (2000)
Volchkov, V. E. et al. Recovery of infectious Ebola virus from complementary DNA: RNA editing of the GP gene and viral cytotoxicity. Science 291, 1965–1969 (2001)
Chan, S. Y., Ma, M. C. & Goldsmith, M. A. Differential induction of cellular detachment by envelope glycoproteins of Marburg and Ebola (Zaire) viruses. J. Gen. Virol. 81, 2155–2159 (2000)
Takada, A. et al. Downregulation of β1 integrins by Ebola virus glycoprotein: implication for virus entry. Virology 278, 20–26 (2000)
Xu, L. et al. Immunization for Ebola virus infection. Nature Med. 4, 37–42 (1998)
Jahrling, P. B. et al. Evaluation of immune globulin and recombinant interferon-α2b for treatment of experimental Ebola virus infections. J. Infect. Dis. 179, S224–S234 (1999)
Parren, P. W., Geisbert, T. W., Maruyama, T., Jahrling, P. B. & Burton, D. R. Pre- and postexposure prophylaxis of Ebola virus infection in an animal model by passive transfer of a neutralizing human antibody. J. Virol. 76, 6408–6412 (2002)
Gupta, M., Mahanty, S., Bray, M., Ahmed, R. & Rollin, P. E. Passive transfer of antibodies protects immunocompetent and immunodeficient mice against lethal Ebola virus infection without complete inhibition of viral replication. J. Virol. 75, 4649–4654 (2001)
Wilson, J. A. & Hart, M. K. Protection from Ebola virus mediated by cytotoxic T lymphocytes specific for the viral nucleoprotein. J. Virol. 75, 2660–2664 (2001)
World Health Organization. Disease Outbreaks. Ebola Haemorrhagic Fever 〈http://www.who.int/disease-outbreak-news/disease/A98.4.htm〉 (2003).
Walsh, P. D. et al. Catastrophic ape decline in western equatorial Africa. Nature 422, 611–614 (2003)
Aoki, K., Barker, C., Danthinne, X., Imperiale, M. J. & Nabel, G. J. Efficient generation of recombinant adenoviral vectors by Cre-lox recombination in vitro. Mol. Med. 5, 224–231 (1999)
Ohno, T. et al. Gene therapy for vascular smooth muscle cell proliferation after arterial injury. Science 265, 781–784 (1994)
Ksiazek, T. G. et al. Enzyme immunosorbent assay for Ebola virus antigens in tissues of infected primates. J. Clin. Microbiol. 30, 947–950 (1992)
Jahrling, P. B. et al. Passive immunization of Ebola virus-infected cynomolgus monkeys with immunoglobulin from hyperimmune horses. Arch. Virol. Suppl. 11, 135–140 (1996)
Acknowledgements
We thank T. Suhana and A. Tislerics for help with manuscript preparation; K. Stroud and T. Miller for figure preparation; D. Levy for helping to define conditions for intracellular cytokine analysis; L. Hensley of USAMRIID and members of the Nabel laboratory for discussions; and T. Larsen for pathology support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing financial interests.
Supplementary information
Rights and permissions
About this article
Cite this article
Sullivan, N., Geisbert, T., Geisbert, J. et al. Accelerated vaccination for Ebola virus haemorrhagic fever in non-human primates. Nature 424, 681–684 (2003). https://doi.org/10.1038/nature01876
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/nature01876
This article is cited by
-
Natural history of nonhuman primates after conjunctival exposure to Ebola virus
Scientific Reports (2023)
-
Mapping HLA-A2, -A3 and -B7 supertype-restricted T-cell epitopes in the ebolavirus proteome
BMC Genomics (2018)
-
Adenovirus-prime and baculovirus-boost heterologous immunization achieves sterile protection against malaria sporozoite challenge in a murine model
Scientific Reports (2018)
-
Post-exposure treatments for Ebola and Marburg virus infections
Nature Reviews Drug Discovery (2018)
-
An adenovirus serotype 2-vectored ebolavirus vaccine generates robust antibody and cell-mediated immune responses in mice and rhesus macaques
Emerging Microbes & Infections (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
