
Abstract
Habitat loss and fragmentation remain the greatest threats to the world's biodiversity1. The local extinction of plant species from habitat fragments is common2, although the reasons for this are not fully understood. Fragmentation is known to influence both birth- and death-related processes3, but the disruption of plant reproduction, especially pollination and seed production, is thought to be particularly important4,5. The effects of fragmentation on post-pollination processes such as seed dispersal and germination have rarely been explored experimentally6. Here I show that seeds planted in forest fragments are less likely to germinate than those in continuous forest. This finding can have negative demographic consequences because it reduces the emergence of seedlings.
Similar content being viewed by others

Main
Heliconia acuminata is an Amazonian understorey herb that is found in both the experimentally isolated rainforest fragments and the continuous forest reserves of Brazil's Biological Dynamics of Forest Fragments Project. During the 1998 fruiting season, which lasted from March to May, I collected 1,668 seeds from more than 200 plants in continuous forest. I then distributed these seeds to seven forest fragments (four fragments of one hectare and three of ten hectares) and three continuous forest sites. Seeds from each maternal plant were distributed evenly across sites to avoid confounding potential maternal effects with fragmentation-related effects.
In each site, seeds were planted every 10 metres along five 100-metre transects. At each location on the transect, a seed was placed on the ground, placed in a plastic cup planted into the ground and filled with local soil (preventing removal of seeds by ants), and placed in a cup buried in the same way but covered with a fine mesh (to prevent all seed predation and accumulation of leaf litter). Seeds were checked each month to see whether they had germinated. One year after planting, I compared the proportion of emerging seedlings in each treatment in each site by using two-way analysis of variance (ANOVA)7.
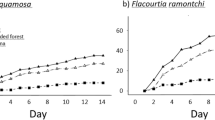
Seeds planted in continuous forest were between three and seven times more likely to germinate than those in forest fragments (P<0.0001; Fig. 1). Seeds in fragments suffered from a variety of edge effects, including hotter, drier conditions and increased light penetration8,9, all of which can affect the cues required for germination. I also found that seeds protected with mesh from leaf-litter accumulation and seed predation were more likely to germinate (although not significantly so), with the improvement being more marked in fragments than in forest sites (2.4- to 4.2-fold compared with 1.8- to 1.9-fold; Fig. 1). This asymmetrical improvement indicates that the greater accumulation of leaf litter in fragments (resulting from slower decomposition8) could be partly responsible for preventing germination as seeds become buried. Higher seed predation in fragments is an unlikely explanation, as predation is extremely low in all sites (E.M.B., unpublished data) and germination rates were similar for seeds placed on the ground and protected from predation in plastic cups.

Differences in the proportion germinating in each habitat and treatment were assessed by ANOVA. The square root of the percentage germinating in each plot was arcsine-transformed, and habitat type and treatment were treated as fixed effects. The effect of habitat type was highly significant (F1,24=22.623, P<0.0001), but that of planting treatment was not (F2,24=2.336, P=0.118). The planting treatment×habitat type interaction term was also nonsignificant (F2,24=0.028, P=0.973). Bars with different letters are significantly different from each other.
The results could be explained by the adaptation of seeds to conditions in the area from which they were collected, with seeds failing to germinate in fragments because of a change in location, rather than as a direct result of fragmentation. However, this mechanism seems unlikely unless seeds are adapted to very broad conditions, such as ‘continuous forest’. Seeds were collected from an area of more than 3.5 km2, and one site of continuous forest was about 20 km from the collection area. When compared with other continuous forest sites, this site never had the lowest germination frequency for any treatment. Even if local adaptation were responsible for the observed pattern, the demographic consequences for Heliconia populations in these fragments would be the same. Populations in fragments have few flowering individuals (E.M.B. and W. J. Kress, unpublished data), so most seeds for these populations probably come from outside the fragments.
Seeds from tropical rainforest plants rarely survive in seed banks for more than a year because they are highly susceptible to fungal pathogens, seed predation and burial under leaf litter10. My results therefore represent the outcome of a population's entire reproductive effort for a year. Although germination success is likely to vary from year to year, my results indicate that seeds dispersed into rainforest fragments are less likely to germinate than those in continuous forest.
Furthermore, my estimates of the effect of fragmentation on seed germination are probably conservative, as plants reproducing in forest fragments may be inbred and suffer from reduced heterozygosity11. If inbred seeds are less likely to germinate, these genetic effects could further reduce the recruitment of seedlings into forest fragment populations, helping to explain why plant populations in habitat fragments often fail to persist in the long term.
References
Wilcove, D. S., Rothstein, D., Dubow, J., Philips, A. & Losos, E. Bioscience 48, 607–615 (1998).
Turner, I. M., Chua, K. S., Ong, J. S. Y., Soong, B. C. & Tan, H. T. W. Conserv. Biol. 10, 1229–1244 (1995).
Jules, E. S. Ecology 79, 1645–1656 (1998).
Bond, W. J. in Extinction Rates (eds Lawton, J. H. & May, R. M.) 131–146 (Oxford Univ. Press, New York, 1995).
Aizen, M. A. & Feinsinger, P. Ecology 75, 330–351 (1994).
Wong, T. C. M., Sodhi, N. S. & Turner, I. M. Biol. Conserv. 85, 97–104 (1998).
Sokal, R. R. & Rohlf, F. J. Biometry (Freeman, New York, 1995).
Didham, R. K. Oecologia 116, 397–406 (1998).
Kapos, V., Wandelli, E., Camargo, J. L. & Ganade, G. in Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities (eds Laurance, W. F. & Bierregaard, R. O. Jr) 33–44 (Univ. Chicago Press, 1997).
Vázquez-Yanes, C. & Orozco-Segovia, A. Annu. Rev. Ecol. Syst. 24, 69–87 (1993).
Young, A., Boyle, T. & Brown, T. Trends Ecol. Evol. 11, 413–418 (1996).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bruna, E. Seed germination in rainforest fragments. Nature 402, 139 (1999). https://doi.org/10.1038/45963
Issue Date:
DOI: https://doi.org/10.1038/45963
This article is cited by
-
Seed high-temperature sensitivity and germination ecology in intermediate seeds of three species from Xishuangbanna, tropical China
Plant Ecology (2023)
-
The dynamic matrix predicts population response to long-term experimental forest fragmentation
Landscape Ecology (2022)
-
Influence of the edge-core gradient on the scorpion assemblage (Arachnida, Scorpiones) in a Brazilian Atlantic forest
International Journal of Tropical Insect Science (2021)
-
Viability of recalcitrant Araucaria angustifolia seeds in storage and in a soil seed bank
Journal of Forestry Research (2020)
-
Edge effects and human disturbance influence soil physical and chemical properties in Sacred Church Forests in Ethiopia
Plant and Soil (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
