
Abstract
Native soil carbon (C) can be lost in response to fresh C inputs, a phenomenon observed for decades yet still not understood. Using dual-stable isotope probing, we show that changes in the diversity and composition of two functional bacterial groups occur with this ‘priming’ effect. A single-substrate pulse suppressed native soil C loss and reduced bacterial diversity, whereas repeated substrate pulses stimulated native soil C loss and increased diversity. Increased diversity after repeated C amendments contrasts with resource competition theory, and may be explained by increased predation as evidenced by a decrease in bacterial 16S rRNA gene copies. Our results suggest that biodiversity and composition of the soil microbial community change in concert with its functioning, with consequences for native soil C stability.
Similar content being viewed by others


Main
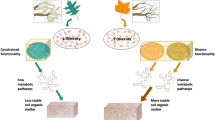
Substrate inputs can stimulate decomposition of native soil organic carbon (SOC; Kuzyakov et al., 2000), a phenomenon known as the ‘priming effect’ (Kuzyakov, 2010), and is considered large enough to influence ecosystem C balance (Wieder et al., 2013). Two functionally distinct groups of microorganisms are postulated to mediate priming: one that grows rapidly utilizing labile C, and one that grows slowly, breaking down recalcitrant SOC (Fontaine et al., 2003; Blagodatskaya et al., 2007). However, distinguishing these groups is technically challenging. Here, we used dual-stable isotope probing with 13C-glucose and 18O-water to identify bacteria in these two groups growing in response to single and repeated pulses of glucose. Organisms that utilize labile C for growth assimilate both 13C-glucose and 18O-water into their DNA, whereas organisms that grow using SOC incorporate only 18O-water. Differential isotope incorporation leads to a range of DNA densities separable through isopycnic centrifugation, which can then be characterized by sequencing (Radajewski et al., 2000).
We sequenced fragments of bacterial 16S rRNA genes following single and repeated glucose pulses. We hypothesized that the single pulse of labile C would stimulate growth of opportunistic organisms, thus immobilizing nutrients and suppressing growth and diversity of the SOC-utilizing community, decreasing SOC decomposition (negative priming), a response analogous to that observed in plant communities in response to chronic N additions (Tilman, 1987; Clark and Tilman, 2008). We hypothesized that multiple glucose additions would stimulate growth of a more diverse bacterial community, including more native SOC-utilizing organisms that possess enzymes to decompose recalcitrant compounds, causing positive priming (Fontaine et al., 2003; Kuzyakov, 2010).
Soil from a ponderosa pine ecosystem was amended weekly for 7 weeks with 500 μg C-glucose per gram soil (2.65 atom % 13C) in 100 μl deionized water or with 100 μl deionized water (n=5). Measurements of δ13C–CO2 and [CO2] enabled the partitioning of CO2 into that derived from added glucose or from native SOC (CSOC):

where Ctotal is CO2–C from glucose-amended samples, δtotal is the δ13C–CO2 from glucose-amended samples, δglucose is the δ13C of the added glucose and δSOC is the δ13C–CO2 evolved from the non-amended samples. Priming was calculated as the difference between SOC oxidation of the amended and non-amended samples. With this approach, any evolved CO2 carrying the 13C signature of the added glucose is considered respiration of glucose, including 13C-labeled biomass and metabolites derived from prior glucose additions. Thus, this approach quantifies priming as the oxidation of SOC present at the beginning of the experiment, consistent with many other studies of priming (Cheng et al., 2003; De Graaff et al., 2010).
In a parallel incubation for dual-stable isotope probing, the repeated-pulse samples received unlabeled glucose (500 μg C-glucose per gram soil) for 6 weeks while the non-amended and single-pulse samples received sterile deionized water. In week 7, samples received one of four isotope treatments (n=3): 97 atom % H2 18O (non-amended soil), 99 atom % 13C-glucose and 97 atom % H2 18O (single- and repeated-pulse soil), 12C-glucose and 97 atom % H2 18O (repeated-pulse soil) or 12C-glucose and H2 16O (repeated-pulse soil). After incubating for 7 days, soil was frozen at −40 °C. DNA was extracted, separated through isopycnic centrifugation, and two density ranges were sequenced for the bacterial 16S rRNA gene (Supplementary Figure 1): 1.731–1.746 g ml−1 (hereafter called the SOC-utilizing community) and 1.759–1.774 g ml−1 (hereafter called the glucose-utilizing community).
Amplicons of the V3–V6 16S rRNA region were bar coded with broad-coverage fusion PCR primers and pooled before sequencing on a Genome Sequencer FLX instrument. These sequence data have been submitted to the GenBank database under accession number SRP043371. Data were checked for chimeras (Edgar et al., 2011), demultiplexed and quality checked (Caporaso et al., 2010). Taxonomy was assigned to genus at the ⩾80% bootstrap confidence level (Cole et al., 2009).
We used the Shannon’s diversity index (H′), commonly used in microbial systems (Fierer and Jackson, 2006), to assess changes in microbial diversity. Analysis of variance was used to compare the amount of DNA within densities between isotope treatments (Supplementary Figure 2) and to test the effects of the treatments on the Shannon’s diversity (Figure 2) and Pielou’s evenness (Supplementary Figure 3) of the active bacterial communities, with post hoc Student’s t-tests, α=0.05. PRIMER 6 and PERMANOVA were used to create the nonmetric multidimensional scaling ordination and to compare bacterial communities between glucose treatments and the two sequenced density ranges.
The single pulse of glucose suppressed SOC oxidation, whereas repeated pulses increased SOC oxidation (Figure 1). Few experiments to date have examined priming in response to repeated substrate amendments (Hamer and Marschner, 2005; Qiao et al., 2014), even though in nature soil receives repeated substrate pulses from litterfall and rhizodeposition. Our results demonstrate the dynamic response of SOC decomposition to repeated labile C inputs.

Weekly priming rates calculated as the difference in SOC respired between glucose-amended and non-amended soil (n=5).
Dual-stable isotope probing was able to separate the growing bacteria into two groups with distinct DNA densities (P<0.001, PERMANOVA; Figure 3a), indicating differential uptake of 13C-glucose and 18O-water. In response to the initial glucose addition, the diversity of the growing glucose- and SOC-utilizing bacterial communities declined compared with the non-amended community (P<0.001, t-tests; Figure 2), driven by a strong decrease in evenness (Supplementary Figure 3). In the SOC-utilizing community, where DNA was labeled with 18O only, the relative abundance of Bacillus increased 4.9-fold compared with the non-amended control to constitute 31.6% of the community (Figure 3b). Bacillus survives well under low-nutrient conditions (Panikov, 1995), and is able to synthesize a suite of extracellular enzymes capable of degrading complex substrates (Priest, 1977), traits that are conducive for using SOC for growth. In the glucose-utilizing community, where DNA was labeled with both 13C and 18O, Arthrobacter increased 67.7-fold relative to the non-amended control to constitute 75.5% of the growing bacteria (Figure 3b). In culture experiments, Arthrobacter can rapidly take up and store glucose for later use (Panikov, 1995) and here we find it dominating the high-density DNA fractions, signifying that it is using the labeled glucose to grow. The increased biomass of Arthrobacter may have resulted in greater resource competition, thus reducing the diversity of the growing community, as is frequently found in plant communities (Bakelaar and Odum, 1978; Clark and Tilman, 2008).

Shannon's diversity index (H′) of the non-amended, single-pulse, and repeated-pulse treatments (n=3) in the SOC- (mid-density) and glucose-utilizing (high-density) communities. Treatments with the same letter are not significantly different from each other (Student's t, α=0.05).

(a) Nonmetric multidimensional scaling ordination showing differences in growing bacterial communities at the genus taxonomic level in the SOC-utilizing (mid-density; open symbols) and glucose-utilizing (high-density; closed symbols) groups of non-amended (Δ), single-pulse (○) and repeated-pulse (□) treatments (n=3). (b) Pie charts of genera in the SOC- and glucose-utilizing communities of the single- and repeated-pulse treatments (n=3). Genera with relative abundances >5% are listed in the figure legend.
After repeated glucose amendments, the diversity of the growing community recovered to non-amendment levels (Figure 2) without strongly dominant organisms (Figure 3b and Supplementary Figure 3). The higher diversity found after repeated glucose pulses may be explained by trophic interactions where predators graze on prey populations that have been enlarged by resource addition, suppressing competition between prey species and causing secondary mobilization of nutrients (Clarholm, 1985). The decrease in total bacterial 16S rRNA gene copies in the repeated-pulse—compared with the single-pulse—treatment (Supplementary Figure 4) supports predation as a potential mechanism explaining the observed diversity increase after repeated glucose pulses.
The recovery of diversity after repeated glucose pulses contrasts with resource competition theory (Tilman, 1987). When chronic additions of a limiting resource are applied, species diversity and evenness typically decrease (Bakelaar and Odum, 1978; Clark and Tilman, 2008) because competitive organisms become dominant. We observed this after the single glucose pulse, but not after repeated pulses. This diversity response may be the result of community shifts facilitated by short bacterial life cycles and the tens to hundreds of generations expected during the 7-week incubation (Behera and Wagner, 1974). In contrast, systems on which most ecological theory is based (for example, plants) might achieve perhaps 20 generations in a multi-decadal field experiment (Bakelaar and Odum, 1978; Clark and Tilman, 2008). With more generations, more community dynamics can occur, including increased resource cascades in which extracellular enzymes, metabolites or lysed cells of one functional group increase substrates for another (Blagodatskaya and Kuzyakov, 2008). Our results highlight the opportunity to test ecological theories in microbial ecosystems (Prosser et al., 2007), particularly as the short life cycles of microbes makes them well suited for pursuing ecological questions in an evolutionary framework (Jessup et al., 2004).
The priming effect is ubiquitous, yet its drivers remain elusive. Our results suggest that changes in the diversity and composition of the growing bacterial community contribute to priming, and thus that ecosystem properties such as soil C storage may be sensitive to soil microbial biodiversity.
Accession codes
References
Bakelaar RG, Odum EP . (1978). Community and population level responses to fertilization in an old-field ecosystem. Ecology 59: 660–665.
Behera B, Wagner GH . (1974). Microbial growth rate in glucose-amended soil. Soil Sci Soc Amer J 38: 591–594.
Blagodatskaya E, Blagodatsky S, Anderson T, Kuzyakov Y . (2007). Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl Soil Ecol 37: 95–105.
Blagodatskaya I, Kuzyakov Y . (2008). Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review. Biol Fertil Soils 45: 115–131.
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7: 335–336.
Cheng W, Johnson D, Fu S . (2003). Rhizosphere effects on decomposition: controls of plant species, phenology, and fertilization. Soil Sci Soc Am J 67: 1418–1427.
Clarholm M . (1985). Interactions of bacteria, protozoa and plants leading to mineralization of soil nitrogen. Soil Biol Biochem 17: 181–187.
Clark CM, Tilman D . (2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451: 712–715.
Cole JR, Wang Q, Cardenas E, Fish J, Chai B, Farris RJ et al. (2009). The Ribosomal Database Project: improved alignments and new tools for rRNA analysis. Nucleic Acids Res 37: D141–D145.
De Graaff M-A, Classen AT, Castro HF, Schadt CW . (2010). Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol 188: 1055–1064.
Edgar RC, Haas BJ, Clemente JC, Christopher Q, Knight R . (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27: 2194–2200.
Fierer N, Jackson R . (2006). The diversity and biogeography of soil bacterial communities. Proc Natl Acad Sci USA 103: 626–631.
Fontaine S, Mariotti A, Abbadie L . (2003). The priming effect of organic matter: a question of microbial competition? Soil Biol Biochem 35: 837–843.
Hamer U, Marschner B . (2005). Priming effects in soils after combined and repeated substrate additions. Geoderma 128: 38–51.
Jessup CM, Kassen R, Forde SE, Kerr B, Buckling A, Rainey PB et al. (2004). Big questions, small worlds: microbial model systems in ecology. Trends Ecol Evol 19: 189–197.
Kuzyakov Y, Friedel J, Stahr K . (2000). Review of mechanisms and quantification of priming effects. Soil Biol Biochem 32: 1485–1498.
Kuzyakov Y . (2010). Priming effects: Interactions between living and dead organic matter. Soil Biol Biochem 42: 1363–1371.
Panikov NS . (1995) Microbial Growth Kinetics. Chapman & Hall: London, UK.
Priest FG . (1977). Extracellular enzyme synthesis in the genus. Bacillus Bacteriol Rev 41: 711–753.
Prosser JI, Bohannan JM, Curtis TP, Ellis RJ, Firestone MK, Freckleton RP et al. (2007). The role of ecological theory in microbial ecology. Nat Rev Microbiol 5: 384–392.
Qiao N, Schaefer D, Blagodatskaya E, Zou X, Xu X, Kuzyakov Y . (2014). Labile carbon retention compensates for CO2 released by priming in forest soils. Global Change Biol 20: 1943–1954.
Radajewski S, Ineson P, Parekh NR, Murrell JC . (2000). Stable-isotope probing as a tool in microbial ecology. Nature 403: 646–649.
Tilman D . (1987). Secondary succession and the pattern of plant dominance along experimental nitrogen gradients. Ecol Monogr 57: 189–214.
Wieder WR, Bonan GB, Allison S . (2013). Global soil carbon projections are improved by modelling microbial processes. Nat Clim Change 3: 909–912.
Acknowledgements
We wish to thank Jim Prosser and three anonymous reviewers for their constructive comments. This material is based upon work supported by the National Science Foundation under grant no.s DEB-1241094 and DBI-1126840.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on The ISME Journal website
Rights and permissions
About this article

Cite this article
Mau, R., Liu, C., Aziz, M. et al. Linking soil bacterial biodiversity and soil carbon stability. ISME J 9, 1477–1480 (2015). https://doi.org/10.1038/ismej.2014.205
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2014.205
This article is cited by
-
Temperature fluctuation promotes the thermal adaptation of soil microbial respiration
Nature Ecology & Evolution (2023)
-
Contrasting priming effect intensities and drivers in single and repeated glucose additions to a forest soil receiving long-term N fertilization
Biology and Fertility of Soils (2023)
-
Stability of Bacterial Network Enhances Nutrient Content in Apple Trees
Journal of Soil Science and Plant Nutrition (2022)
-
Soil nutrient variability mediates the effects of erosion on soil microbial communities: results from a modified topsoil removal method in an agricultural field in Yunnan plateau, China
Environmental Science and Pollution Research (2022)
-
Soil properties resulting in superior maize yields upon climate warming
Agronomy for Sustainable Development (2022)
