
Abstract
Animals that modify their physical environment by foraging in the soil can have dramatic effects on ecosystem functions and processes. We compared bacterial and fungal communities in the foraging pits created by bilbies and burrowing bettongs with undisturbed surface soils dominated by biocrusts. Bacterial communities were characterized by Actinobacteria and Alphaproteobacteria, and fungal communities by Lecanoromycetes and Archaeosporomycetes. The composition of bacterial or fungal communities was not observed to vary between loamy or sandy soils. There were no differences in richness of either bacterial or fungal operational taxonomic units (OTUs) in the soil of young or old foraging pits, or undisturbed soils. Although the bacterial assemblage did not vary among the three microsites, the composition of fungi in undisturbed soils was significantly different from that in old or young foraging pits. Network analysis indicated that a greater number of correlations between bacterial OTUs occurred in undisturbed soils and old pits, whereas a greater number of correlations between fungal OTUs occurred in undisturbed soils. Our study suggests that digging by soil-disturbing animals is likely to create successional shifts in soil microbial and fungal communities, leading to functional shifts associated with the decomposition of organic matter and the fixation of nitrogen. Given the primacy of organic matter decomposition in arid and semi-arid environments, the loss of native soil-foraging animals is likely to impair the ability of these systems to maintain key ecosystem processes such as the mineralization of nitrogen and the breakdown of organic matter, and to recover from disturbance.
Similar content being viewed by others


Introduction
Australia has suffered one of the highest rates of global mammal extinctions over the past 200 years since European settlement (Woinarski et al., 2012). Losses have been most pronounced in the critical weight range (35–5500 g) mammals, which were once common over large areas of continental Australia (Johnson, 2006). The loss of these animals, or the contraction of their ranges, has been attributed to multiple causes associated with European settlement and pastoral practices such as altered fire regimes, overgrazing by livestock, competition with exotic pests including the European rabbit (Oryctolagus cuniculus) and predation by introduced species such as the domestic cat (Felis catus) and the red fox (Vulpes vulpes) (Johnson, 2006). Two species that have suffered substantial range restrictions are the greater bilby (Macrotis lagotis) and the burrowing bettong (Bettongia lesueur). Recent attempts have been made to reintroduce these animals into predator-proof exclosures within their former range in an effort to re-establish viable populations (James and Eldridge, 2007).
Many of Australia’s locally extinct animals forage extensively in the soil for seeds, bulbs, invertebrates and fungi (Robley et al., 2001; James et al., 2011; Eldridge et al., 2012). Foraging disturbs the soil surface and breaks up the surface crust (biocrust), altering rates of water infiltration, and creating small pits and depressions that trap water, soil, organic matter and seed (James et al., 2009). These pits develop into patches of higher nutrients, with greater concentrations of plant-available nitrogen and carbon than the surrounding soil matrix (James et al., 2010) and often a different vegetation community (Lavelle et al., 2006). Studies worldwide have shown that modification of the abiotic environment by these animals, a process referred to as ecosystem engineering (sensu Jones et al., 1994), alters energy flows and resource availability, increases species richness, diversity and productivity, through niche construction, ultimately controlling the availability and distribution of resources to other organisms (for example, Whitford and Kay, 1999; Jones et al., 2010).
An important process moderated by soil-disturbing animals in arid environments is the decomposition of organic matter. Litter and organic matter in these systems are spatially and temporally variable, and is often concentrated within the foraging pits of animals (James and Eldridge, 2007). Litter is a source of carbon, nitrogen and other trace elements, and provides habitat for a range of micro- and macro-invertebrates involved in the decomposition of organic matter (Haslem et al., 2011). Litter falling into pits comes into close contact with soil, where it is held in situ more effectively than if it remained on the soil surface where it is subject to removal by wind and water (Whitford, 2002). Together with reduced evaporation resulting from lower temperatures in the pits than the undisturbed surface (Eldridge and Mensinga, 2007), this increases the time period over which soil moisture is optimum for microbial decomposition and nutrient mineralization (Steinberger and Whitford, 1983; Jacobsen and Jacobsen, 1998; Whitford, 2002). Litter remaining on the surface, however, is subject to photodegradation (Austin and Vivanco, 2006), potentially reducing the return of carbon to the soil organic pool.
Soil-disturbing animals therefore play an important role in bringing surface-resident organic matter into contact with soil microorganisms. The accumulation of litter in the pits is also likely to exert a strong selective pressure on microorganisms essential for the decomposition process. Given the marked differences in the biotic (litter cover and composition) and abiotic (for example, surface temperature, soil moisture) environments between pits and undisturbed soils, that is, those soils undisturbed by animal activity (for example, Vossbrinck et al., 1979; Wallwork et al., 1985; Eldridge and Mensinga, 2007), we expected that the pits would differ in the composition of soil microorganisms. For example, studies of foraging disturbances constructed by the short-beaked echidna (Tachyglossus aculeatus) indicate a greater diversity and abundance of micro-arthropods and higher rates of microbial respiration in the pits than undisturbed soil (Eldridge and Mensinga, 2007), suggesting that there are differences in the abundance or structure of microbial communities. Over time, pits collect organic matter, and research has shown that pits over about 12 months old have high levels of organic carbon. This compares with young pits (<3 months old), which have relatively low levels of litter and organic matter (D J Eldridge, unpublished data). We would expect pit age to influence microbial community structure, as these old pits (~12 months) would have more time to establish seedlings and accumulate litter and microorganisms that are present on adjacent, undisturbed surfaces. Furthermore, older pits could provide a greater range of different environments, with differences in depth, shape and orientation, and therefore different soil chemistry and organic matter at varying stages of decomposition.
We compared the community structure of soil microbial communities in old and young pits with the undisturbed surface soil on two soil types in an area where bilbies and bettongs have been reintroduced into their former range. Both bilbies and bettongs construct pits while foraging for buried seed, invertebrates and plant roots. The pits of these two species are indistinguishable, and range from cylindrical-shaped excavations about 15-cm wide and up to 20-cm deep to shallow basin-like structures (Eldridge et al., 2012). Pits are constructed only once, and unlike cache pits of heteromyid rodents (Geluso, 2005), are rarely reworked. Because pits vary in depth and shape, and are constructed in soils of different texture, they provide a range of different physical environments that influences the trapping and retention of litter and the breakdown of organic matter.
We hypothesized that the microbial community in pit soils would support more microorganisms commonly associated with decomposing litter. Conversely, we expected that the microbial community composition in undisturbed soils would support a community dominated by cyanobacteria, given the extensive cover of biocrusts on the soil surface. We used microbial network analysis to examine the structure of microbial communities, particularly in relation to resilience and reactivity (Ruiz-Moreno et al., 2006; Bissett et al., 2013). Examination of microbial networks improves our understanding of why undisturbed soils might be resistant to nutrient amendment, how microbial community structure is altered following pit construction, and how digging promotes nutrient enrichment within these microsites (James et al., 2009).
Materials and methods
The study area
Our study was undertaken within the Australian Wildlife Conservancy’s Scotia Sanctuary in south-western, New South Wales, Australia (33°43’S, 143°02’E) where locally extinct bilbies and bettongs have been released into predator-proof exclosures. Soil samples were collected from two systems; (i) mallee (Eucalyptus spp.) west-east-trending dunes of Quaternary alluvium characterized by calcareous and siliceous sands (Rudosols) and (ii) the inter-dunal swales and plains extending to these dunes, which are up to 500-m wide, comprised mainly of loamy, calcareous soils (Calcarosols). The vegetation on the dunes is moderately dense mallee (Eucalyptus socialis, E. dumosa) and the plains are dominated by open mallee woodland with scattered belah (Casuarina pauper) and sugarwood (Myoporum platycarpum), and a variable cover of shrubs such as punty bush (Senna artemisioides), hopbush (Dodonaea viscosa), turpentine (Eremophila sturtii), pinbush wattle (Acacia burkittii) and assorted bluebushes (Maireana spp.), depending on whether trees had been removed. Shrubs covered about 50% of the area of the plains. The climate in the area is semi-arid, with cool winters (mean⩽17 °C) and hot summers (mean 30 °C). Rainfall is highly spatially and temporally variable and averages 243 mm per year. Rainfall is evenly distributed between the six warmer months and the six cooler months.
Field sampling
The location, size, depth and age of all foraging pits constructed by bilbies and bettongs have been monitored at 36 large sites at the Scotia Sanctuary since 2007. Because sites were visited every 3 months, we were able to calculate the relative age of particular pits. In October 2009, we collected soil samples from six sites: three on sandy dunes and three on loamy plains. At each of the six sites, we sampled three microsites: (i) young foraging pits, that is, pits constructed since the previous measurements (<3 months old), (ii) old foraging pits, that is, pits older than 12 months and (iii) undisturbed non-pit surface soils at least 3 m from any pit. At each of the six sites, we sampled each microsite at 10 locations. For the young pits, soil was removed from the uppermost 10 mm layer of the soil surface or from the base of the pits after removing any existing organic material. Biocrust was not removed from the soil prior to sampling. Approximately 5 g of soil was collected with a sterilized spatula. The material from the 10 locations was then bulked and stored on ice before being transported back to the laboratory. The same procedure was used to collect samples from old pits and undisturbed surfaces. This resulted in a database of 18 bulked samples (3 replicate sites of 2 soil types × 3 microsites).
Molecular analysis
Environmental DNA was isolated from 500 mg of soil using the FASTDNA Spin Kit for Soil (MP Bio Laboratories, Inc., Carlsbad, CA, USA) according to the manufacturer’s instructions and stored at −80 °C until use. DNA was quantified using a NanoDrop ND-1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and the quality checked by PCR amplification of the 16 S rRNA gene using the primer pair 27 f/519r (Weisburg, 1991). Bacterial and fungal specific tag-encoded FLX amplicon pyrosequencing of each sample was carried out using the primers 27 f/519r and funSSUF/funSSUR respectively (Lucero, 2011) on a Roche GS-FLX Titanium at the Research and Testing Laboratory (Lubbock, TX, USA). Sequence reads were analyzed using MOTHUR v1.22 (www.mothur.org) software package (Schloss et al., 2009). Initial quality processing of 454 sequence reads was performed using the mothur implementation of PyroNoise (Quince et al., 2011) using default settings. Sequences containing <200 bp, containing ambiguous bases and homopolymers longer than 8 bp in length were removed. The remaining sequences were aligned to either the bacterial or fungal alignments of the SILVA release 102 reference alignment. Chimeric sequences were identified and removed using the mothur implementation of uchime (Edgar et al., 2011). The taxonomic identity of each unique sequence was determined by comparison against the SILVA release 102 reference database. Taxonomic assignment was made at each level, given a bootstrap value greater than 80, using the RDP classifier (Wang et al., 2007). Sequences that failed to be classified at the phylum level or were classified as either Mitochondria, Archaea or Eukaryota/Prokaryota in the respective datasets, were removed. Sub-sampling was performed at a level of 400 sequences per sample for the bacterial dataset and 1300 sequences per sample for the fungal dataset. Implementation of this process resulted in the exclusion of a bacterial young loam soil sample and bacterial young sand soil sample, as these samples contained fewer than the 400 sequences required. To ensure a balanced design across the bacterial dataset, the corresponding samples were subsequently excluded from the bacterial old pit soil and bacterial surface pit soil sets (2 replicate sites of 2 soil types × 3 microsites. Uncorrected pairwise distances were calculated between sequence reads with the final clustering of operational taxonomic units (OTUs) performed at a 0.03 distance threshold using the average neighbor algorithm (Schloss and Westcott, 2011). The identity of each OTU defined at 0.03 distance threshold was obtained from the consensus of each sequence within that OTU at a confidence threshold of 80. From these data, two individual data matrices were generated, one for bacteria and one for fungi, each matrix containing every OTU and the number of reads assigned to it from each sample. In this instance, the relative proportion of each OTU was used as a proxy for abundance, as absolute abundance measures were not obtained.
Statistical analysis
We used permutation multivariate analysis of variance (PERMANOVA; Anderson et al., 2008) to examine differences in the composition of a data matrix of 2500 bacterial OTUs, defined at 0.03 distance threshold, and a data matrix of 5895 fungal OTUs, defined at 0.03 distance threshold, in relation to microsite (undisturbed soils, young pits, old pits) and soil type (loam, sand). Relative abundance data were used after resampling, to ensure an equivalent number of sequences. The first stratum of this analysis considered soil type and the second stratum microsite and its interaction with soil type. Pairwise a posteriori comparisons were made, where necessary, using a multivariate analogue of the t statistic, the probability levels being obtained by permutation. We tested for differences in richness and diversity of taxa with a mixed-model ANOVA with the same structure as the PERMANOVA analysis. Richness and diversity data were checked for homogeneity of variance (Levene’s test) and normality using diagnostic tests but no transformations were needed. For all analyses, significant differences between means were examined using Fisher’s Protected Least Significant Difference test. The procedure was repeated for the fungal data.
The degree of association of OTUs with respect to microsite was measured with Indicator Species Analysis in R (De Cáceres et al., 2012) using a data matrix consisting of 2500 bacterial OTUs and 5895 fungal OTUs. Indicator values combine information on relative abundance and frequency of species, and the indicator value is maximal (IV=100%) when all individuals of a given species are restricted to a particular microsite (for example, old pit), and all samples from the particular microsite contain an occurrence of that species. Data (at the OTU level) were randomized among the treatments and a Monte Carlo randomization procedure performed with 1000 iterations to determine the statistical significance of the indicator values.
The degree of association of OTUs with respect to one another within each microsite was measured using the Pearson’s correlation coefficient (r). Bacterial and fungal OTU tables, defined at 0.03 distance threshold, were separated on the basis of microsite, and then reduced by removing any OTUs that did not occur across at least 75% of available samples. A Pearson’s r score and P-value were calculated pairwise for each bacterial OTU using the rcor.test algorithm, available from the ltm package (available from https://cran.r-project.org/package=ltm) as implemented in R version 3.0.2. For each correlation, P-values were generated and the false discovery rate was maintained below 5% using the Benjamini-Hochberg procedure (Benjamini and Hochberg, 1995). Visualization of these interactions, incorporating taxonomic, abundance and microsite occurrence information, was made with the freely available Cytoscape package version 2.8.3 (available at: www.cytoscape.org). For each network, topological metrics of connectivity and density were calculated using the network analysis plug-in (Assenov et al., 2008). Networks pre-embedded with sample and OTU-specific information are provided in the Supplementary Material.
Results
Richness of bacterial and fungal taxa
Most bacterial and fungal OTUs occurred at very low abundances, with a substantial number of abundances equal to one. Of the original 2500 bacterial OTUs after resampling, 320 (14%) contributed 50% of total OTU abundances. For fungal OTUs, 525 (9%) of the 5895 OTUs contributed 50% of total fungal abundances. There were no differences in bacterial OTU richness (that is, different number of OTUs) among the different soils (P=0.24; range 238–332 OTUs) or among the three microsites (P=0.47). Similarly, fungal richness did not vary with soil texture (P=0.81; range 397–873) or among the three microsites (P=0.17).
Community composition of bacterial and fungal taxa
Bacterial communities were observed to contain a high proportion of Actinobacteria, and to a lesser extent, Alphaproteobacteria and Acidobacteria (Figure 1a). Cyanobacteria appeared to constitute a large proportion of the bacterial community, particularly in undisturbed soils. Fungal communities were observed to contain a high proportion of Lecanoromycetes, and to a lesser extent, Archaeosporomycetes (Figure 1b).

Relative abundance of major (a) bacterial and (b) fungal taxa within each microsite. Larger circles indicate greater abundance.
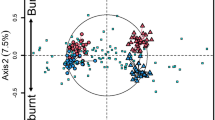
There was no significant difference in the composition of either bacterial or fungal OTUs assemblages between loamy and clay soils (P>0.30). The composition of the bacterial assemblage did not vary among the three microsites (P=0.21; Figure 2a), but there was a significant effect for fungi (Pseudo F2,8=3.08, P(Perm)=0.003). The composition of fungi in undisturbed soils was significantly different from that in old (pairwise t=2.14, P=0.029) or young (t=2.02, P=0.02) pits, but there was no significant differences between old and young pits (P=0.47; Figure 2b).

Multi-dimensional scaling biplot of the first two dimensions of an ordination of a reduced matrix of (a) 280 bacterial OTUs and (b) 135 fungal OTUs. Note the clustering of undisturbed samples for both bacteria and fungi.
Microsite indicators
Six cyanobacterial OTUs (Gp I (3 OTUs), Gp X, Gp VII and an unclassified OTU) were indicators of undisturbed pits, as were the single Asanoa OTU (Actinobacteria), a Segetibacter OTU (Sphingobacteria) and an unclassified alphaproteobacterial OTU. A single Hylangium OTU (Myxobacteria, Deltaproteobacteria), Microvirga OTU (Rhizobiales, Alphaproteobacteria) and a Gp IV actinobacterial OTU were indicators of old pits. A single Rubrobacter OTU (Actinobacteria), Ammoniphilus OTU (Bacilli, Firmicutes) and Actinaurispora OTU (Actinobacteria) were indicators of young pits (Table 1). Overall, fungal taxa were better discriminators of the three microsites, with 20 orders containing 170 OTUs, with indicator values >0.70, and almost exclusively from sub-phylum Pezizomycotina. These included orders Dothideales (genera Columnosphaeria, Delphinella), Chaetothyriales (genus Glyphium), Lecanorales (genera Sphaerophoraceae, Cladoniaceae), Myxotrichaceae (genus Geomyces), Mycocaliciales (Sphinctrina) and Pleosporales (genera Leptosphaeria, Trematosphaeria, Phaeosphaeria). Ten fungal genera (particularly Eupenicillium, Hamigera, Bionectriaceae and an unclassified taxon from the family Bulgaria) were highly indicative (IV>0.81) of young pits. Old pits contained a wide range of different OTUs, with the orders Chaetothyriales, Dothideales, Hypocreales, Lecanorales, Mycocaliciales and Pleosporales having a large number of OTUs that were strongly indicative (IV>69%) of older pits (Table 2).
Network analysis
Within the bacterial networks, the mean number of correlations between OTUs was greater in old pit soils (3.45) than either undisturbed (2.516) or young pit (1.294) soils, consistent with a larger number of OTUs co-occurring across the samples (Table 3). The majority of the associations present in young pit soils were between a small number of alphaproteobacterial and actinobacterial OTUs. Young pit soils returned the lowest values for network metrics of clustering (0), density (0.081) and centralization (0.050). Undisturbed soils and old pit soils were similar in relation to clustering (undisturbed=0.566, old pits=0.547), density (undisturbed=0.084, old pits=0.088) and centralization (undisturbed=0.089, old pits=0.096) (Table 3).
Within the fungal networks, the highest mean number of correlations between OTUs (20.497) was observed in undisturbed soils, where many more OTUs (1814 OTUs) were present across multiple samples than in young (321) or old (485) pit soils. Similar to the bacterial networks, young pit soils returned the lowest values for density (0.009), but were also the most centralized (0.116). Old pit soils were the least clustered (0.472) and the least centralized (0.067). Undisturbed soils were similar to young pit soils in terms of clustering (undisturbed=0.652, young pits=0.647), whereas fungal young and old pit soils were only similar in relation to the mean number of correlations between OTUs.
Discussion
Soil foraging by semi-fossorial animals in arid areas disrupts surface crusts, alters rates of water infiltration, and creates small pits and depressions that trap water, soil, organic matter and seed (James and Eldridge, 2007). We expected to detect substantial differences in the soil microbial community between intact undisturbed soils and recently excavated or older, more established pits in response to differences in plant and litter cover, organic matter decomposition and soil nutrient concentrations. Although we detected some significant differences in the fungal community composition between the soil surface and the pits (described below), there were no discernible differences in the bacterial community and in the fungal community between young and old pits, largely because of the high variability among microsites (Figure 1). Consequently, we undertook an analysis that would test whether the physical variability that is observed in pit soils, in regard to moisture and nutrient trapping, influenced the occurrence of individual species or the manner in which individual species exhibited correlations with one another. Indicator species analysis was implemented to identify specific OTUs that were more strongly associated with a particular microsite type. Critically, indicator species analysis has been previously shown to be suitable for identifying variable taxa where there was no prior assessment, or no significant variation, in the larger community composition (De Cáceres and Legendre, 2009; De Cáceres and Legendre, 2009; De Cáceres et al., 2012). That it were possible to identify species that were statistically indicative of particular microsites when the multivariate (PERMANOVA) analysis was insignificant highlights the fact that there is substantial heterogeneity within microsites, and suggests a level of functional redundancy within microbial taxa that prevents large-scale perturbation of the community despite the loss of species. On the basis of the indicator species (De Cáceres and Legendre, 2009; De Cáceres et al., 2012) and microbial network (Chaffron, 2010; Bissett et al., 2013) analyses, there is sufficient evidence to suggest that pits may be associated with a reduction in autotrophic groups (Figure 1, Tables 1 and 2) that are compensated for by an emergence of taxa capable of decomposing organic material (Tables 1 and 2) and reduced resilience in the microbial communities (Table 3).
Compositional differences between pit and undisturbed soils
Consistent with information from arid soils worldwide, the bacterial community contained high proportions of Actinobacteria and Alphaproteobacteria (Figure 1a) (Yeager et al., 2004; Kuske, 2012). At the community level, we detected no significant differences in bacterial community composition between pits and undisturbed soils (Figure 2). However, consistent with our first hypothesis, filamentous diazotrophic (Cyanobacteria GpI), baeocystous (Cyanobacteria GpVIII) and unicellular (Cyanobacteria GpX) cyanobacteria were found to be indicators of undisturbed soils (Table 1) with a reduction in the observed abundance of cyanobacterial sequence reads when soils were disturbed (Figure 1, Table 1). Cyanobacteria were present in undisturbed soils, however, the presence of these taxa as indicators was reflective of both a decrease in the abundance of cyanobacterial groups and a shift within the morphological and physiological nature of cyanobacteria between undisturbed and pit soils. Among the heterotrophic population, actinobacterial members of the Rubrobacteridae that are pioneers in biological crust formation (Yeager et al., 2004) dominated both undisturbed and pit soils, with a single Rubrobacter OTU, an indicator of young pit soils. In addition to cyanobacterial groups, the Sphingobacterial genus Segetibacter has been previously affiliated with the decomposition of cyanobacteria- and plant-derived phytodetritus (Li et al., 2011).
Fungal communities in undisturbed and pit soils comprised a wide range of saprotrophs, with Lecanoromycetes, the largest class of lichenized fungi, and to a lesser extent, Archaeosporomycetes, comprising about 80% of sequences across the three microsites (Figure 1b). Along with Pezizomycotina, these fungal taxa perform a diverse array of ecological functions including wood and litter decomposition, mycorrhizal associations and lichen symbioses, animal and plant pathogens (Spatafora et al., 2006). Evidence for active recession, or at least competitive inhibition, of microbial groups from the old pits was found, with the insect and plant pathogenic fungi, Delphinella, Leptosphaeria, Trematosphaeria and Columnosphaeria, found almost exclusively in undisturbed and young pit soils. Glomeromycetes, which comprise arbuscular mycorrhizal species, represented about 3% of sequences in young pits and 2% of sequences in old pit and undisturbed soils.
Community development with pit age
Rubrobacter, Ammoniphilus and Actinaurispora were the only bacterial indicators of young pits and likely represent remnants of the sub-surface community. Rubrobacter is a cosmopolitan and abundant taxon in arid zone soils (Yeager et al., 2004). The presence of Ammoniphilus and Actinaurispora in young pit soils is likely due to the deposition of plant material. Amminophilus has been reported as a strictly aerobic oxalotroph utilizing plant- and algae-derived oxalic acid as a sole carbon. Actinaurispora are known plant endophytes, inhabiting Camptotheca acuminata species (Zhu et al., 2012). The family Micromonosporacaeae, to which Actinaurispora belongs, however, has been tentatively correlated with increasing moisture content in arid and semi-arid soils (Bachar et al., 2010), which may contribute to the presence of this species as an indicator of young soils. Trichocomaceae species were the primary fungal indicators of young pits. A single Trichocomaceae species was a key fungal indicator of undisturbed soils, suggesting that fungal communities of young pits contain residual surface taxa prior to the colonization and diversification of fungal communities observed in older pits. Trichocomaceae species are predominantly saprotrophic, have aggressive colonization strategies and a high tolerance to extreme environmental conditions such as soil drying, high temperature and metal toxicity (Houbraken and Samson, 2011). Their presence in young pits could indicate opportunistic colonization of recently disturbed soil.
On the basis of the criteria used to select the microsites, progression of the microbial community from young to old pits occurs over a period of 9–12 months. Over this time, although little change occurred within the microbial community composition between pit stages, a discernable difference was observed between the undisturbed and pit bacterial and fungal communities, irrespective of their age. Microbial richness among microsites, however, remained unchanged. Spore propagule density and arbuscular mycorrhizal fungi diversity are known to decline with increasing tillage associated with agriculture (Brito et al., 2010; Schalamuk et al., 2013). However, this was not reflected in our fungal species richness, which remained unchanged over time. The progressive accumulation of fungal species attached to organic matter and seed in the pits is consistent with the presence of several lichenized lecanoralean genera including Parmeliaceae and Myrangium (Smith, 1948) and the epiphytic melanized taxon Sarcinomyces (Wollenzien et al., 1997). The Lecanorales are predominantly lichen-forming fungi that are mycobionts of the genera Xanthoparmelia, Parmotrema and Xanthoria, which are common corticolous lichens of Callitris glaucophylla trees that occur in the study area (Filson and Rogers, 1979). These taxa are typically found on the soil surface or in the pits on detached plant material. At some sites, we also recorded the vagant lichen Chondropsis semiviridis from within the pits. This lichen, which has no attachment to the soil, moves freely along the surface by wind action (Eldridge and Leys, 1999). Similarly, Cladonia spp., another common soil lichen genus, was found on undisturbed surfaces. Along with the lichen genera Endocarpon and Placidium, it is one of the most common lichens forming biocrusts on stable soils in arid and semi-arid areas (Eldridge and Koen, 1998).
Despite our inability to discriminate between the bacterial community of old and young pits, we recorded three indicator species, Hyalangium and Microvirga, and a Gp IV Acidobacteria. The two proteobacterial species were indicative of the presence of established vascular plants. Hyalangium, belongs to the group of Myxobacteria that uses plant lignin and produces small bioactive molecules. Microvirga has been implicated in nodule formation, facilitating nitrogen-fixing processes within the rhizosphere (Ardley et al., 2012). The occurrence of these groups in old pit soils is likely to enhance nitrogen fixation, presumably to levels greater than those in the undisturbed and young pit soils, and support the growth of vascular plants occurring in these microsites.
Microbial co-occurrence in pit and undisturbed soils
Our analyses thus far indicate that initial disturbance reduces the abundance of key photoautotrophic groups, and that over extended periods of time, capture of organic matter leads to changes in the abundance of some taxa, with increases in those taxa likely reflecting an increased capacity for the assimilation of organic carbon and nitrogen matter.
Resilience is the ability of a system to recover from large disturbances, typically over short time frames. Reactivity, however, is the capacity of a system to respond to small perturbations over extended periods. Under such circumstances, the apparent equilibrium may appear stable, despite moving to a new steady state over long time periods (Neubert et al., 2009). Modularity, defined by the number and size of groups of highly interconnected nodes within a network, is positively correlated with reactivity, and negatively correlated with resilience (Ruiz-Moreno et al., 2006). Analysis of both bacterial and fungal microbial networks revealed stark differences in modularity, reflected in the values of clustering, density and centralization, of microbial co-occurrence networks between undisturbed soils and pit soils at different developmental stages (Table 3). Clustering coefficients and density (network connectivity) scores tending towards a value of 1 indicate a highly modular system whereas those tending towards zero represent the opposite (Bissett et al., 2013). Low values of clustering and density associated with microbial communities from contaminated and reference estuarine sediments indicate historical community 'stress' contributing to functional redundancy and reduced correlations among species (Sun et al., 2013). This was reinforced by marginally lower values, for each of these metrics, in contaminated sediments, with the suggestion that this anthropogenic perturbation has contributed an additional stress.
In the present study, bacterial species–species correlations within the young pit soils were almost nonexistent. A clustering coefficient of zero and a slightly lower density value were consistent with reduced modularity, and an increase in functional redundancy associated with a recent external stress (Sun et al., 2013). In contrast, undisturbed and old pit soils were more consistent with increased modularity, suggesting a lack of functional redundancy, with greater species–species correlations, and increased clustering and density. This suggests to us that the bacterial community present in undisturbed soils and old pit soils are more reactive and less resilient than young pit soils. Within the fungal communities, the number of correlations among species, clustering coefficients and density, and hence modularity, were highest in surface soils and lowest in the old pit soils, suggesting that fungal communities within old pits are less reactive and more resilient. In contrast, the young pit soils exhibit reduced modularity, and increased resilience, suggesting that they are likely to respond to nutrient amendments over the short-term, thereby driving large and dramatic structural changes. This is largely because of the high degree of physical disturbance created when foraging pits are established. Within the old pit soils, the bacterial community has largely regained the modularity observed within the undisturbed soils. The fungal community, however, is apparently more resistant at this stage than in the undisturbed soils, suggesting it is able to continue to drive structural changes in response to events such as litter deposition.
A high level of centralization, as a consequence of the high frequency of centralized nodes, was observed among the fungal community in young pit soils (Bulgariaceae, Myxotrichaeae, Trichocomaceae, Tubeufiaceae) and among the bacterial community in undisturbed (Rubrobacteraceae, Geodematophiliaceae, Bradyrhizobiaceae) and old pit (Rhodobacteraceae, Bradyrhizobiaceae, Geodermatophiliaceae, Beijerinckiaceae, Comamonadaceae, Methylobacteriaceae) soil (Supplementary Information). Centralized nodes have been proposed to represent keystone species, exhibiting a large influence of the ‘information’ transfer throughout the community (Bisssett et al., 2013). It has been speculated that these nodes represent critical control points in the cycling of nutrients within the system (Ruiz-Moreno et al., 2006; Bissett et al., 2013). Thus, it is realistic to suggest that these centralized taxa act to stabilize the microbial community. It should be highlighted that these observations were made in the context of a small number of samples defining each microsite, as well as few sequence reads being available to identify species–species correlations. Our observations between the bacterial and fungal datasets suggest that these metrics are susceptible to sequence depth, and pre-treatment of the data by retaining only semi-ubiquitous (occurring across at least 75% of samples) OTUs. These values may also be influenced by the level of heterogeneity within microsites. Despite this, our analyses of network metrics from the bacterial communities suggested that the community structure of old pit soils reflect that of undisturbed soils. Over the long term, this would tend towards decreased responses to nutrient inputs into these soils. This, however, may be partially offset by frequent deposition of plant matter due to the establishment and growth of vascular plants within old pit soils, and subsequent assimilation of this matter by saprotrophic fungi.
Conclusions
Our study suggests that digging by soil-disturbing animals is likely to create successional shifts in soil microbial and fungal communities, which could account for increases in organic matter of nitrogen in old foraging pits (James et al. 2009). The observed richness of fungal and bacterial OTUs, in undisturbed soils, and young and old pits did not differ, though fewer correlations, and hence an increased resilience, were observed between bacterial OTUs in young pits, and fungal OTUs in young and old pits. This suggests that these communities are more likely to respond over the short term to nutrient amendment, thus promoting nutrient enrichment and contributing to a form of patchiness that is critical for the functioning of arid systems. The action of soil-disturbing animals therefore leads to the development of a mosaic of different patches with a varying complement of microorganisms. Given the wide variety in pit size, depth, substrate and spatial configuration, this differential microbial activity will likely lead to the creation of a mosaic of patches of differing resource availability, analogous to larger surface-resident biotic patches such as hummocks and debris mounds. Our work suggests that microbial community composition and co-occurrence change with physical disturbance during the formation of foraging pits. Given the primacy of organic matter decomposition in arid and semi-arid environments, the loss of native soil-foraging animals from these systems may well impair the ability of these systems to maintain key ecosystem processes and to recover from disturbance.
References
Anderson MJ, Gorley RN, Clarke KR . (2008). PERMANOVA+ for PRIMER: guide to software and statistical methods. PRIMER-E: Plymouth, UK.
Ardley JK, Parker MA, De Meyer SE, Trengove RD, O'Hara GW, Reeve WG et al. (2012). Microvirga lupini sp. nov., Microvirga lotononidis sp. nov. and Microvirga zambiensis sp. nov. are alphaproteobacterial root-nodule bacteria that specifically nodulate and fix nitrogen with geographically and taxonomically separate legume hosts. Int J System Evol Microb 62: 2579–2588.
Assenov Y, Ramirez F, Schelhorn SE, Lengauer T, Albrecht M . (2008). Computing topological parameters of biological networks. Bioinformatics 24: 282–284.
Austin AT, Vivanco L . (2006). Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 442: 555–558.
Bachar A, Al-Ashhab A, Soares MIM, Sklarz MY, Angel R, Ungar ED, Gillor O . (2010). Soil microbial abundance and diversity along a low precipitation gradient. Microb Ecol 60: 453–461.
Benjamini Y, Hochberg Y . (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Royal Stat Soc Ser B 57: 289–300.
Bissett A, Brown MV, Siciliano SD, Thrall PH . (2013). Microbial community responses to anthropogenically induced environmental change: towards a systems approach. Ecol Lett 16: 128–139.
Brito I, Goss MJ, de Carvalho M, Chatagnier O, van Tuinen D . (2010). Impact of tillage system on arbuscular mycorrhiza fungal communities in the soil under Mediterranean conditions. Soil Till Res 121: 63–67.
Chaffron S, Rehrauer H, Pernthaler J, von Mering C . (2010). A global network of coexisting microbes from environmental and whole-genome sequence data. Genome Res 20: 947–959.
De Cáceres M, Legendre P . (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90: 3566–3574.
De Cáceres M, Legendre P, Wiser SK, Brotons L . (2012). Using species combinations in indicator analyses. Method Ecol Evol 3: 973–982.
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R . (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27: 2194–2200.
Eldridge DJ, Mensinga A . (2007). Foraging pits of the Short-Beaked Echidna (Tachyglossus aculeatus) as small-scale patches in a semi-arid Australian box woodland. Soil Biol Biochem 39: 1055–1065.
Eldridge DJ, Leys JF . (1999). Wind dispersal of the vagant lichen Chondropsis semiviridisin semi-arid eastern Australia. Aust J Bot 47: 157–164.
Eldridge DJ, Koen TB . (1998). Cover and floristics of microphytic soil crusts in relation to indices of landscape health. Plant Ecol 137: 101–114.
Eldridge DJ, Koen TB, Huang N, Killgore A, Whitford WG . (2012). Animal foraging as a mechanism for sediment movement and soil nutrient development: evidence from the semi-arid Australian woodlands and the Chihuahuan Desert. Geomorph 157/158: 131–141.
Filson RB, Rogers RW . (1979). Lichens of South Australia. Govemment Printer: South Australia.
Geluso K . (2005). Benefits of small-sized caches for scatter-hoarding rodents: influence of cache size, depth, and soil moisture. J Mammal 86: 1186–1192.
Haslem A, Kelly LT, Nimmo DG, Watson SJ, Kenny SA, Taylor RS et al. (2011). Habitat or fuel? Implications of long-term, post-fire dynamics for the development of key resources for fauna and fire. J Appl Ecol 48: 247–256.
Houbraken J, Samson RA . (2011). Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Studies Mycol 70: 1–51.
Jacobsen KM, Jacobsen PJ . (1998). Rainfall regulates decomposition of buried cellulose in the Namib Desert. J Arid Environ 38: 571–583.
James AI, Eldridge DJ . (2007). Reintroduction of fossorial native mammals and potential impacts on ecosystem processes in an Australian desert landscape. Biol Conserv 138: 351–359.
James AI, Eldridge DJ, Hill BM . (2009). Foraging animals create fertile patches in an Australian desert shrubland. Ecography 32: 723–732.
James AI, Eldridge DJ, Moseby KE . (2010). Foraging pits, litter, and plant germination in an arid shrubland. J Arid Environ 74: 516–520.
James AI, Eldridge DJ, Koen TB, Moseby KE . (2011). Can the invasive European rabbit (Oryctolagus cuniculus) assume the soil engineering role of locally-extinct natives? Biol Invas 13: 3027–3038.
Johnson C . (2006) Australia’s mammal extinctions: a 50000 year history. Cambridge University Press: Melbourne.
Jones CG, Lawton JH, Shachak M . (1994). Organisms as ecosystem engineers. Oikos 69: 373–386.
Jones CG, Gutierrez JL, Byers JE, Crooks JA, Lambrinos JG, Talley TS . (2010). A framework for understanding physical ecosystem engineering by organisms. Oikos 119: 1862–1869.
Kuske CR, Yeager CM, Johnson S, Ticknor LO, Belnap J . (2012). Response and resilience of soil biocrust bacterial communities to chronic physical disturbance in arid shrublands. ISME J 6: 886–897.
Lavelle P, Daecans T, Aubert M, Barot S, Blouin M, Bureau F et al. (2006). Soil invertebrates and ecosystem services. Eur J Soil Biol 42: S3–S15.
Li H, Xing P, Chen M, Bian Y, Wu QL . (2011). Short-term bacterial community composition dynamics in response to accumulation and breakdown of Microcystis blooms. Water Res 45: 1702–1710.
Lucero ME, Unc A, Cooke P, Dowd S, Sun S . (2011). Endophyte microbiome diversity in micropropagated Atriplex canescens and Atriplex torreyi var griffithsii. PLoS One 6: e17693.
Neubert MG, Caswell H, Solow AR . (2009). Detecting reactivity. Ecology 90: 2683–2688.
Quince C, Lanzen A, Davenport RJ, Turnbaugh PJ . (2011). Removing noise from pyrosequenced amplicons. BMC Bioinf 12: 38.
Robley AJ, Short J, Bradley S . (2001). Dietary overlap between the burrowing bettong and the European rabbit in semi-arid coastal Western Australia. Wildl Res 28: 341–349.
Ruiz-Moreno D, Pascual M, Riolo R . (2006). Exploring network space with genetic algorithms: modularity, resilience and reactivity. In: Pascual M, Dunne JA (eds), Ecological networks: linking structure to dynamics in food webs. Oxford University Press: UK, pp 187–208.
Schalamuk S, Velázquez S, Cabello M . (2013). Dynamics of arbuscular mycorrhizal fungi spore populations and their variability under contrasting tillage systems in wheat at different phonological stages. Biol Agric Hortic 29: 38–45.
Schloss PD, Westcott SL, Ryanbin T, Hall JR, Hartmann M, Hollister EB et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Micro 75: 7537–7541.
Schloss PD, Westcott SL . (2011). Assessing and improving methods used in operational taxonomic unit-based approaches for 16 S rRNA gene sequence analysis. Appl Environ Micro 77: 3219–3226.
Smith HD . (1948). Biological control of citrus insects. In: Webber HJ, Batchelor LD (eds), The citrus industry The production of the crop Vol. II. California Press: USA, pp 597–626.
Spatafora JW, Sung GH, Johnson D, Hesse C, O'Rourke B, Serdani M et al. (2006). A five-gene phylogeny of Pezizomycotina. Mycologia 98: 1018–1028.
Steinberger Y, Whitford WG . (1983). The contribution of rodents to decomposition processes in a desert ecosystem. J Arid Environ 6: 177–182.
Sun MY, Dafforn KA, Johnston EL, Brown MV . (2013). Core sediment bacteria drive community response to anthropogenic contamination over multiple environmental gradients. Environ Micro 15: 2517–2531.
Vossbrinck CR, Coleman DC, Woolley TA . (1979). Abiotic and biotic factors in litter decomposition in a semiarid grassland. Ecology 60: 265–271.
Wallwork JA, Kamill BM, Whitford WG . (1985). Distribution and diversity patterns of soil mites and other microarthropods in a Chihuahuan desert site. J Arid Environ 9: 215–231.
Wang Q, Garrity GM, Tiedje JM, Cole JR . (2007). Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Micro 73: 5261–5267.
Weisburg WG, Barns SM, Pelletier Da, Lane DJ . (1991). 16 S ribosomal DNA amplification for phylogenetic study. J Bacteriol 173: 697–703.
Whitford WG . (2002) Ecology of Desert Systems. Elsevier Science: London.
Whitford WG, Kay FR . (1999). Biopedturbation by desert mammals: a review. J Arid Environ 41: 203–230.
Woinarski JCZ, Burbidge AA, Harrison PL . (2012) The Action Plan for Australian Mammals, 2012 CSIRO Publishing: Melbourne, Australia.
Wollenzien U, Hoog GS, Krumbein WE, Uijthf JMJ . (1997). Sarcinomycespetricola, a new microcolonial fungus from marble in the Mediteranean Basin. Antone van Leeuwenhoek 71: 281–288.
Yeager CM, Kornosky JL, Housman DC, Grote EE, Belnap J, Kuske CR . (2004). Diazotrophic community structure and function in two successional stages of biological soil crusts from the Colorado Plateau and Chihuahuan Desert. Appl Environ Micro 70: 973–983.
Zhu WY, Zhao LX, Zhao GZ, Duan XW, Qin S, Li J et al. (2012). Plantactinospora endophytica sp. nov., an actinomycete isolated from Camptotheca acuminata Decne., reclassification of Actinaurispora siamensis as Plantactinospora siamensis comb. nov. and emended descriptions of the genus Plantactinospora and Plantactinospora mayteni. Int J Syst Evol Micro 62: 2435–2442.
Acknowledgements
We thank Ivan Wong for field assistance and Samantha Travers for comments on an earlier draft of the manuscript. We acknowledge the considerable financial, logistic and technical support provided by the Australian Wildlife Conservancy through their Scotia Sanctuary. This research was supported by the Australian Government under ARC LP0882630.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on The ISME Journal website
Supplementary information
Rights and permissions
About this article

Cite this article
Eldridge, D., Woodhouse, J., Curlevski, N. et al. Soil-foraging animals alter the composition and co-occurrence of microbial communities in a desert shrubland. ISME J 9, 2671–2681 (2015). https://doi.org/10.1038/ismej.2015.70
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2015.70
This article is cited by
-
Recurring heavy rainfall resulting in degraded-upgraded phases in soil microbial networks that are reflected in soil functioning
Soil Ecology Letters (2023)
-
Foraging pit location provides valuable insights into critical habitat requirements of soil engineers
Landscape Ecology (2023)
-
A reintroduced ecosystem engineer provides a germination niche for native plant species
Biodiversity and Conservation (2020)
-
Digging mammal reintroductions reduce termite biomass and alter assemblage composition along an aridity gradient
Oecologia (2019)
-
Keystone taxa as drivers of microbiome structure and functioning
Nature Reviews Microbiology (2018)
