
Abstract
This study employed microsatellite loci to analyse outcrossing rate and pollen dispersal in Japanese red pine (Pinus densiflora) in an isolated stand. The average offspring outcrossing rate for 29 cones was 0.955. Significant differences in outcrossing rates between offspring groups on individual branches that extended in different directions at different heights were not detected. Male parents of 874 offspring collected from the maternal tree were assessed by exclusion using polymorphisms at three microsatellite loci. Paternity analysis indicated that at least 31% of the offspring were fertilized by pollen from trees outside the stand. The average distance of pollen migration within the study stand was 68 m, with a maximum value of 325 m. There was excess mating with nearby P. densiflora trees, of which only a few were predominant pollen donors. In addition, a weakly directional bias in P. densiflora pollination was also detected in the study stand, suggesting that female strobili on a branch of the maternal tree were more easily fertilized by pollen from trees in that direction.
Similar content being viewed by others


Introduction
Japanese red pine (Pinus densiflora Sieb. & Zucc.) is one of the most widely distributed Japanese conifers. This species grows naturally on a wide variety of sites, extending from the foot of Mt. Tarumai in Hokkaido (30°15′ N), the northernmost island in Japan, to the southern part of Yaku Island (42°40′ N), and from sea level to an altitude of 2290 m (Hayashi, 1952). Extensive mortality, resulting from damage caused by pinewood nematodes (Bursaphelenchus xylophilus), has gradually increased in southwestern Japan since the 1940s. Knowledge of the mating processes and patterns of natural effective pollen movement may be indispensable for successful pine forest protection and regeneration, although these have not yet been reported in Japanese red pine.
Patterns of mating and gene flow via pollen dispersal within populations strongly affect genetic variation and the rate and direction of microevolutionary change (Schuster & Mitton, 2000). However, pollen dispersal based on measuring physical pollen movements using traps (Greenwood, 1986) or dyes (Linhart et al., 1987; Campbell, 1991) may not always reflect actual fertilization and gene flow among local populations (for review, see Dow & Ashley, 1998a, b; Streiff et al., 1999).
Recently, approaches using molecular markers have been used to estimate effective gene flow by comparing the segregation of genetic markers in the parental and offspring cohorts. In particular, there has been a rapid increase in studies using microsatellite markers to estimate mating patterns and pollen dispersal in natural tree populations (Dow & Ashley, 1996; Dawson et al., 1997, 1998a; Dayanandan et al., 1999; Streiff et al., 1999; Isagi et al., 2000; Ueno et al., 2000). Such markers allow estimation of allele frequencies, and a high exclusion probability for paternity assignment, owing to their codominant inheritance and high polymorphism.
In this study, highly polymorphic microsatellite markers were used to estimate outcrossing rate and effective pollen donors for offspring produced on individual branches of a single Japanese red pine growing in an isolated stand.
Materials and methods
Study site
The study site was in the University of Tokyo’s Experimental Station at Tanashi, located in suburban Tokyo. The Experimental Station occupies 9.12 ha and is surrounded by residential areas. Only a few scattered individual pines grow in parks and gardens around the experimental station. There was no prevailing wind from April to May 1998 during the flowering period of P. densiflora.
The experimental station contains two groups of Japanese red pine (Fig. 1). The first group consists of 101 trees that are about 70 years old and over 15 m high. These trees arose from natural regeneration and are scattered throughout the experimental station. The second group comprises 53 individuals about 20 years old and 10 m high, clustered in the northern part of the Experimental Station (Plot A, Fig. 1), that were planted from the seeds of trees growing in the experimental station. The positions of the 70-year-old trees were obtained from a survey distributed by the station; those of the 20-year-old trees in Plot A were measured using a survey laser instrument (Criterion 300, Laser Technology, Englewood, Co., USA).

Map of the Pinus densiflora study stand. Closed and open circles represent trees that were approximately 70 and 20-year-old, respectively. M is the maternal tree.
Sample collection and preparation
For the outcrossing and paternity analysis, a maternal tree of 26 cm d.b.h. (diameter at breast height) and 12 m height, branching above 3 m, located to the west of most of the trees in Plot A was chosen (Fig. 1). In October 1999, two or three apparently undamaged mature cones were collected from branches that extended in different directions at different heights on the maternal tree (Table 1). Seeds were collected from each cone and stored at 4°C for 40 days. In order to facilitate germination, seeds were soaked in distilled water at 4°C overnight and then sown in individual pots containing an autoclaved (120°C, 2 h) mixture (1:1 v/v) of Tanashi nursery soil (black sand loam) and Shibanome soil (volcanic sand) in a greenhouse (25°C day: 23°C night). After 30 days, all the needles were removed from each seedling and DNA was immediately extracted. If more than 34 seedlings were available from a cone, then 34 seedlings were randomly chosen for analysis. Fresh needles were also collected from all 154 adult trees in the experimental station.
DNA extraction and microsatellite analysis
Crude genomic DNA was extracted from fresh P. densiflora needles using a modified CTAB method. About 100 mg material was homogenized for at least 45 s in 700 μL 2 × ΧΤΑΒ solution using a FastPrep™ FP120 machine (Savant, USA), and incubated at 65°C for 1 h. DNA was isolated using chloroform–isoamyl alcohol (24:1) extraction, precipitation in isopropanol, and washing in 75% ethanol. Extracted DNA was dissolved in 100 μL Tris-EDTA (TE) buffer. For paternity analysis, four microsatellite PCR primer pairs (Pde3, Pde5, Pde7, Pde14), developed by Lian et al. (2000), were used to score genotypes. The PCR reaction conditions were the same as those described previously (Lian et al., 2000). The reaction products were electrophoresed on a 6% Long Ranger sequencing gel (FMC BioProducts Co., ME, USA) using a SQ-5500 sequencer (Hitachi Co., Tokyo). Electrophoretic patterns were analysed with FRAGLYS v.2 software (Hitachi Electronics Engineering Co., Tokyo).
Data analysis
For each locus, allele frequencies were determined, and Nei’s (1987) method was employed to calculate expected heterozygosity (HE). Differences in allele frequency distribution between adults and the offspring produced on individual branches of the maternal tree were tested using the Kolmogorov–Smirnov test (Morin et al., 1994). The multilocus outcrossing rate (tm) was estimated using a maximum likelihood method (Shaw et al., 1981). Paternity was assessed by simple exclusion, based on multilocus genotypes (Dow & Ashley, 1996; Schnabel, 1998; Streiff et al., 1999). Possible paternal alleles at every locus, inferred by subtracting the maternal alleles from offspring alleles, were compared with those of adult trees; adults that did not share the possible paternal alleles at least at one locus were excluded as not being possible male parents of the offspring.
The correlations between fertilization frequency of offspring in the maternal tree by pollen from an individual paternal tree and distance of the paternal tree from the maternal tree were statistically analysed. The paternal trees were classified according to their distance from the maternal tree at 10-m intervals. The average frequency of offspring fertilized by paternal trees that were included in each distance class, was plotted against distances from the maternal tree. The correlation was estimated using a negative exponential model.
In order to investigate fertilization bias caused by the directions of paternal trees to the mother tree, the surrounding area of the maternal tree was divided into four quadrants, northeastern (NE), southeastern (SE), southwestern (SW), and northwestern (NW) quadrants, as shown in Fig. 1. The frequency of offspring fertilized by paternal trees in each quadrant were calculated for offspring in cones on branches which were extended in each direction. The frequency distribution of paternity of trees in the four quadrants was compared between branches extending in different directions.
Results
Allele frequency
No alleles at locus Pde7 of the maternal tree were amplified in 50% of the progeny, indicating there was a common null allele at this locus. Thus, locus Pde7 was excluded from further analysis.
The total number of alleles per locus ranged from 24 to 37 in adult trees and from 7 to 24 in offspring produced on individual branches of the maternal tree (Table 2). At all loci, several alleles appearing in the offspring genotypes were absent from the adults. The heterozygosities of individual loci ranged from 0.762 to 0.915 in adult trees and from 0.602 to 0.936 in the offspring produced on individual branches of the maternal tree (Table 2).
The differences in allele frequency distribution between each offspring group sampled from individual branches and adult trees and between the offspring groups were tested using the Kolmogorov–Smirnov test. The allele frequencies at loci Pde5 and Pde14 in adult trees were significantly different from those in every offspring group (P < 0.05). Although the allele frequencies at locus Pde3 in most offspring groups were significantly different from those in adult trees (P < 0.05), groups S1 and NW2 did not show different allele frequencies from adult trees. No significant differences among offspring groups were found at any locus.
Outcrossing rate
Outcrossing rates for offspring from individual cones varied from 0.777 to over 1 (mean 0.955); of the 29 cones investigated, the offspring outcrossing rates in 11 cones (38%) were over 1 (Fig. 2). F-statistics did not show significant differences among the outcrossing rates of offspring on different branches of the maternal tree, except for those in the northwestern group (Table 3). On northwestern branches of the maternal tree, the outcrossing rate of offspring from the upper cones (NW1) was significantly higher than that from the middle (NW2) and lower (NW3) cones (P < 0.05).

Frequency distribution of multilocus outcrossing rates among offspring sampled from individual Pinus densiflora cones. Cones were grouped according to multilocus outcrossing rates of offspring within them at 0.03 intervals.
The germination frequency of seeds from individual cones ranged from 56.3 to 85.3%, and there was a significant positive correlation between germination frequency and outcrossing rate (r=0.407, d.f.=29, P < 0.05; Fig. 3).

Correlation between germination frequency and multilocus outcrossing rate of offspring sampled from individual Pinus densiflora cones. The asterisk (*) indicates that the coefficient was significant at P < 0.05.
Paternity analysis
By simple exclusion analysis, at least 38 of the 154 adult trees within the study stand did not fertilize any offspring in the maternal tree. Twelve of these 38 trees grow in Plot A, which contains 83 adult trees. For 604 out of the 874 offspring analysed, possible male parents were identified as adult trees within the study stand. Of these 604 offspring, 403 had a single possible male parent and 201 had two or more possible male parents. Simple exclusion analysis indicated that 56 were produced by self-fertilization. In contrast, 270 offspring had no compatible male parents within the study stand. Therefore, their male parents were outside the study stand.
The pollen migration distances for the 347 outcrossing offspring for which single male parents were identified within the Experimental Station, ranged from 6 to 325 m, with an average of 68 ± 72 m (SD) (Fig. 4). There was a significant negative correlation between the number of offspring fertilized by an individual tree and the distance between the paternal and maternal trees in the study stand.

Correlation between offspring fertilization frequency in the maternal Pinus densiflora tree by pollen from given paternal trees and distance between the parents. Double asterisks (**) indicate that the coefficient was significant at P < 0.01.
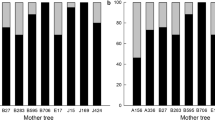
The distribution of paternal trees that fertilized offspring produced on individual branches of the maternal tree was significantly biased according to branch direction (P < 0.05, χ2 test) (Figs 5 and 6). Offspring produced on southern branches of the maternal tree were fertilized relatively more often by pollen from adult trees south-east and north-east of the maternal tree. Offspring on the northeastern and northwestern branches were fertilized relatively more often by pollen from trees to the north-east. Pollen donors for offspring produced on the south-western branches of the maternal tree were mostly to the south-east. There was a conspicuous tendency for pollen from a small number of trees to fertilize the offspring, i.e. eight out of 154 individuals fertilized 42% of the offspring of the maternal tree. Trees 2 and 33 fertilized 9.8 and 9.2% of the total offspring, respectively (Fig. 5). Tree 33, which was north-east of the maternal tree, fertilized 20% of the offspring on the northeastern branches, 10% on the southern branches, 5.1% on the northwestern branches, and 4.2% on the southwestern branches.

Location of pollen donors within Plot A for offspring produced on branches of maternal Pinus densiflora tree that extended to south-west, south, north-east, and north-west. •, c. 70-year-old trees; ○, c. 20-year-old trees. Line width connecting a tree with the maternal tree represents the number of offspring that were inferred to be fertilized by pollen from that individual. The narrowest and broadest lines represent 1 and 17 offspring, respectively.

Frequency distribution of offspring fertilized by paternal trees in three quadrants, i.e. north-east (NE), south-east (SE) and south-west (SW) within offspring from seeds borne on the maternal Pinus densiflora tree branches that extend south-west, south, north-east or north-west. The surrounding area of the maternal tree was divided into four quadrants: northeastern (NE), southeastern (SE), southwestern (SW), and northwestern (NW). There were no trees in NW quadrant. The frequency distribution of paternity of offspring from each directional group of branches was plotted against the three quadrants.
Within the study stand, no trees were located to the north-west of the maternal tree. The fertilization frequency of offspring on branches of the maternal tree pointing in different directions by pollen from outside the stand varied. The highest levels of fertilization by pollen from outside trees were detected in offspring produced on the southwestern and northwestern branches and were 38% and 36%, respectively. The lowest rate, 24%, occurred on the southern branches.
Discussion
Outcrossing rate of P. densiflora
The multilocus outcrossing rate of P. densiflora estimated in this study stand averaged 0.955, and was similar to that of other pine species: 0.862 for P. washoensis (Mitton et al., 1997), 0.987 for P. sylvestris (Burczyk, 1998), and 0.944 for P. ponderosa (Sorensen, 1994). This value is higher than that of most broad-leafed species, such as Helicteres brevispira (tm=0.478–0.676) (France schinelli & Bawa, 2000), Eucalyptus rhodantha (tm=0.59–0.65), and Cavanillesia platanifolia (tm=0.213–0.569) (Murawski, 1995). Mitton (1992) suggested that conifers generally produce seeds by outcrossing, and the high outcrossing rate of P. densiflora is consistent with this hypothesis.
In this study, the offspring outcrossing rate from each cone produced on individual branches of the maternal tree did not depend on branch position, i.e. the height or direction of the branch bearing the cone, and varied considerably among cones from 0.77 to over 1.00. From 11 out of 29 cones, all offspring were produced by outcrossing. Although no comparison between cones of offspring outcrossing rates on an individual pine has previously been reported, investigators have reported dependence and independence of the outcrossing rate on the height of branches or position of trees within a population. Franklin (1971) reported that the outcrossing rate in P. taeda seeds depended on sampling height. Shaw & Allard (1982) also reported a similar dependence on tree position within Douglas-fir populations along roadsides or the edges of natural breaks. On the contrary, almost no dependence of the outcrossing rate on tree position was observed in a young Douglas-fir grafted orchard (Shaw & Allard, 1982), a low-density P. banksiana stand with trees of various heights (Cheliak et al., 1985), or an even-aged, natural P. contorta stand (Perry & Dancik, 1986). Comparing the growth forms of P. contorta and P. ponderosa, Sorensen (1994) suggested that denser branching of P. contorta than P. ponderosa corresponds to its lower outcrossing rate. The maternal P. densiflora tree in our study was about 20 years old and sparsely branched. This shape might have permitted pollen from surrounding P. densiflora trees free access to its female strobili, resulting in the high outcrossing rate, as well as eliminating the effect of branch position on the outcrossing rate.
There was a significant positive correlation between germination frequency and outcrossing rate, from the correlation formula, under complete outcrossing and selfing, germination frequencies of P. densiflora seeds are estimated as 75.0% and 24.4%, respectively. Using these two germination frequencies, we estimated the average outcrossing rate of seeds to be 0.874. This indicates that the outcrossing rate (0.955) estimated from the seedlings produced from seeds might be an overestimate. Inbreeding depressions of seed viability, germination and growth tend to be severe for conifers (Mitton et al., 1997), eliminating inbred individuals between the seed and adult life stages. Therefore, the above inferences also imply that selfed seeds in P. densiflora may be at a selective disadvantage relative to outcrossed seeds.
Paternity analysis of offspring produced on individual branches in a P. densiflora tree
Polymorphism analysis using codominant markers enables inference of the pollen dispersal pattern. The paternity exclusion analysis of P. densiflora indicated that pollen from trees outside the study stand produced 270 (31%) of the 874 analysed offspring of the maternal tree. Based on the position of paternal trees within the study stand, the average distance of pollen migration within the study stand was 68 m with a maximum value of 325 m (Fig. 4). These results correspond to the long-distance pollen migration reported in P. flexilis (Schuster & Mitton, 2000), P. sylvestris (Harju & Nikkanen, 1996), and Quercus macrocarpa (Dow & Ashley, 1996), and confirm the previous suggestion that gene flow in conifers may occur across long distances (Levin & Kerster, 1974; Schuster & Mitton, 2000).
Fertilization of offspring of the maternal tree with pollen from a paternal tree seemed to be correlated with the distance between the trees. P. densiflora pollen is windborne, and pollen density may be greater nearer the paternal tree. A significant negative correlation was found between the fertilization frequency by pollen from a given tree and its distance from the maternal tree (Fig. 4). This suggests that pollen density around female strobili might be one of the determining factors of fertilization in P. densiflora. However, this finding is not in agreement with the results for P. flexilis (Schuster & Mitton, 2000) and P. ponderosa (Latta et al., 1998), which do not show an excess of near-neighbour matings. Pollen scattering by wind is influenced greatly both by meteorological conditions and tree growth form. Therefore, the correlation between fertilization frequencies and the distance between the parent trees may be labile, and condition dependent.
A directional bias of fertilization on individual branches of the maternal P. densiflora tree by trees located in different directions was seen with the direction of branch extension (Fig. 5). Within the study stand, no trees were located north-west of the maternal tree. The northwestern and southwestern branches of the maternal tree had higher levels of outside pollination. Moreover, the potential paternal trees distributed north of the maternal tree contributed more pollen to the northern branches of the maternal tree (Fig. 5). Female strobili on a given side of the maternal tree might be pollinated more easily by adult trees on the same side of the maternal tree. These results further confirm that the pollen density around female strobili might be an important factor for fertilization in P. densiflora. To our knowledge, pollination bias according to branch direction has not yet been investigated. To date, most investigations on the direction bias of pollination have focused on the level of the whole tree (Dow & Ashley, 1998a; Streiff et al., 1999). Streiff et al. (1999) indicated that although fertilization in seven of the 13 maternal trees investigated was directionally biased in a mixed oak stand of Quercus robur and Q. petraea, it was not biased in the other six maternal trees. Tauber (1977) pointed out that modelling pollen dispersal is complex because of numerous sources of variation, such as wind direction and velocity, meteorological conditions, and local disturbances. Therefore, the directional bias of fertilization found on individual branches of the maternal P. densiflora tree might also vary with conditions.
The pollen that fertilized most of the offspring of the maternal tree was derived from a relatively small number of paternal trees. Dow & Ashley (1998a) reported a similar result, in that much of the pollen that effectively fertilized seeds of Q. macrocarpa came from a small number of trees. As discussed above, if pollen density around female strobili primarily determines the fertilization frequencies, this means that a small number of trees in the population produced much more pollen than other trees, or had a phenology compatible with the maternal tree.
In conclusion, P. densiflora seeds were produced mainly by outcrossing, although the offspring fertilization frequency by the pollen of an individual paternal tree was somewhat correlated with the distance and direction of the paternal tree from the female strobili. Paternity tended to be confined to a small portion of the trees surrounding the maternal tree. The characteristics of pollen dispersal in P. densiflora demonstrated in the present study might be common to every windborne species, e.g. other Pinus species.
References
Burczyk, J. (1998). Mating system variation in a Scots pine clonal seed orchard. Silvae Genet, 47: 155–158.
Campbell, D. R. (1991). Comparing pollen dispersal and gene flow in a natural population. Evolution, 45: 1965–1968.
Cheliak, W. M., Dancik, B. P., Morgan, K., Yeh, F. C. H. and Strobeck, C. (1985). Temporal variation of the mating system in a natural population of jack pine. Genetics, 109: 569–584.
Dawson, I. R., Waugh, R., Simons, A. J. and Powell, W. (1997). Simple sequence repeats provide a direct estimate of pollen-mediated gene dispersal in the tropical tree Gliricidia sepium. Mol Ecol, 6: 179–183.
Dayanandan, Y., Dole, J., Bawa, K. and Kesseli, R. (1999). Population structure delineated with microsatellite markers in fragmented populations of a tropical tree, Carapa guianensis (Meliaceae). Mol Ecol, 8: 1585–1592, 10.1046/j.1365-294x.1999.00735.x.
Dow, B. D. and Ashley, M. V. (1996). Microsatellite analysis of seed dispersal and parentage of saplings in bur oak, Quercus macrocarpa. Mol Ecol, 5: 615–627.
Dow, B. D. and Ashley, M. V. (1998a). High levels of gene flow in bur oak revealed by paternity analysis using microsatellites. J Hered, 89: 62–70.
Dow, B. D. and Ashley, M. V. (1998b). Factors influencing male mating success in bur oak, Quercus macrocarpa. New Forests, 15: 161–180, 10.1023/a:1006557904751.
Franceschinelli, E. V. and Bawa, K. S. (2000). The effect of ecological factors on the mating system of a South American shrub species (Helicteres brevispira). Heredity, 84: 116–123, 10.1046/j.1365-2540.2000.00636.x.
Franklin, E. C. (1971). Estimates of frequency of natural selfing and inbreeding coefficients in loblolly pine. Silvae Genet, 20: 194–195.
Greenwood, M. S. (1986). Gene exchange in loblolly pine: the relation between pollination mechanism, female receptivity and pollen availability. Am J Bot, 73: 1443–1451.
Harju, A. M. and Nikkanen, T. (1996). Reproductive success of orchard and nonorchard pollens during different stages of pollen shedding in a Scots pine seed orchard. Can J For Res, 26: 1096–1102.
Hayashi, Y. (1952). The natural distribution of important trees, indigenous to Japan. Conifers Report 2: Bull Gov Forest Exp St, Tokyo, 55: 241–242.
Isagi, Y., Kanazashi, T., Suzuki, W., Tanaka, H. and Abe, T. (2000). Microsatellite analysis of the regeneration process of Magnolia obovata Thunb. Heredity, 84: 143–151, 10.1046/j.1365-2540.2000.00642.x.
Latta, R. G., Linhart, Y. B., Fleck, D. and Elliot, M. (1998). Direct and indirect estimates of seed versus pollen movement within a population of ponderosa pine. Evolution, 52: 61–67.
Levin, D. A. and Kerster, H. W. (1974). Gene flow in seed plants. Evol Biol, 7: 139–220.
Lian, C., Miwa, M. and Hogetsu, T. (2000). Isolation and characterization of microsatellite loci from the Japanese red pine. Mol Ecol, 9: 1186–1188, 10.1046/j.1365-294x.2000.00954-10.x.
Linhart, Y. B., Busby, W. H., Beach, J. H. and Feinsinger, P. (1987). Forager behavior, pollen dispersal and inbreeding in two species of hummingbird-pollinated plants. Evolution, 41: 679–682.
Mitton, J. B. (1992). The dynamic mating systems of conifers. New Forests, 6: 197–216.
Mitton, J. B., Latta, R. G. and Rehfeldt, G. E. (1997). The pattern of inbreeding in Washoe pine and survival of inbred progeny under optimal environmental conditions. Silvae Genet, 46: 215–218.
Morin, P. A., Moore, J. J., Chakraborty, R. J. I. N. L., Goodall, J. and Woodruff, D. S. (1994). Kin selection, social structure, gene flow, and the evolution of flowering time. Science, 265: 1193–1201.
Murawski, D. A. (1995). Reproductive biology and genetics of tropical trees from a canopy perspective. In: Loman, M. and Nadkarni, N. (eds) Forest Canopy, pp. 457–493. Academic Press, New York.
Nei, M. (1987). Molecular Evolutionary Genetics. Columbia University Press, New York.
Perry, D. J. and Dancik, B. P. (1986). Mating system dynamics of lodgepole pine in Alberta, Canada. Silvae Genet, 35: 190–195.
Schnabel, A. (1998). Parentage analysis in plants: mating systems, gene flow, and relative fertilities. In: Carvalho, G. R. (ed.) Advances in Molecular Ecology, pp. 173–189. IOS Press, Oxford.
Schuster, W. S. F. and Mitton, J. B. (2000). Paternity and gene dispersal in limber pine (Pinus flexilis James). Heredity, 84: 348–361, 10.1046/j.1365-2540.2000.00684.x.
Shaw, D. V. and Allard, R. W. (1982). Estimation of outcrossing rates in Douglas-fir using isozyme markers. Theor Appl Genet, 62: 113–120.
Shaw, D. V., Kahler, A. L. and Allard, R. W. (1981). A multilocus estimator of mating system parameters in plant populations. Proc Natl Acad Sci USA, 78: 1298–1302.
Sorensen, F. C. (1994). Frequency of seedlings from natural self-fertilization in Pacific Northwest ponderosa pine (Pinus ponderosa Dougl. ex Laws.). Silvae Genet, 43: 100–108.
Streiff, R., Ducousso, A., Lexer, C., Steinkelllner, H., Gloessl, J. and Kremer, K. (1999). Pollen dispersal inferred from paternity analysis in a mixed oak stand of Quercus robur L. & Q. petraea (Matt.) Liebl. Mol Ecol, 8: 831–841, 10.1046/j.1365-294x.1999.00637.x.
Tauber, M. (1977). Investigations of aerial pollen transport in a forested area. Dansk Bot Arkiv, 32: 1–121.
Ueno, S., Tomaru, N., Yoshimaru, H., Manabe, T. and Yamamoto, S. (2000). Genetic structure of Camellia japonica L. in an old-growth evergreen forest, Tsushima, Japan. Mol Ecol, 9: 647–656, 10.1046/j.1365-294x.2000.00891.x.
Acknowledgements
This work was supported in part by a grant from PROBRAIN and Grants-in-Aid from the Ministry of Education, Science, Sports and Culture of Japan. We thank the staff of Experimental Station at Tanashi, the University of Tokyo, for providing the tree map reference and their help in sampling.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lian, C., Miwa, M. & Hogetsu, T. Outcrossing and paternity analysis of Pinus densiflora (Japanese red pine) by microsatellite polymorphism. Heredity 87, 88–98 (2001). https://doi.org/10.1046/j.1365-2540.2001.00913.x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1046/j.1365-2540.2001.00913.x
Keywords
This article is cited by
-
Population genetics information for the regional conservation of a tropical seagrass, Enhalus acoroides, around the Guimaras Strait, Philippines
Conservation Genetics (2017)
-
Development and transferability of two multiplexes nSSR in Scots pine (Pinus sylvestris L.)
Journal of Forestry Research (2015)
-
Fine-scale distribution of ectomycorrhizal fungi colonizing Tsuga diversifolia seedlings growing on rocks in a subalpine Abies veitchii forest
Mycorrhiza (2014)
-
A hybrid swarm population of Pinus densiflora × P. sylvestris inferred from sequence analysis of chloroplast DNA and morphological characters
Journal of Forestry Research (2013)
-
Extensive selfing in an endangered population of Pinus parviflora var. parviflora (Pinaceae) in the Boso Hills, Japan
Tree Genetics & Genomes (2013)
