
Abstract
Populations of the insect-pollinated geophytes Narcissus assoanus and N. dubius (Amaryllidaceae) are commonly dimorphic for stigma height. An extensive survey of populations of the two species in SW France revealed a wide range of style-morph frequencies, particularly populations with significantly more long-styled than short-styled plants. Here we employ experimental and theoretical approaches to investigate potential selective mechanisms governing the variation in style-morph frequencies. Controlled pollination of both species demonstrated that N. assoanus is moderately self-sterile whereas N. dubius is highly self-compatible. Both intra- and intermorph crosses of N. assoanus were equally fertile, indicating that the species does not exhibit heteromorphic incompatibility. Estimates of female fertility (fruit- and seed-set) and multilocus estimates of outcrossing using allozyme markers provided no evidence of morph-specific differences in maternal components of reproductive success in natural populations of the two species. This result suggested that differences between the morphs in male fertility may be largely responsible for the observed morph-ratio variation. To investigate this hypothesis we developed a mating model that incorporates the genetics of stigma-height dimorphism and contrasting rates of assortative and disassortative mating in the style morphs. Simulation results demonstrated that stigma-height dimorphism will always be maintained when levels of disassortative mating are greater than assortative mating, and that the observed L-biased populations in Narcissus spp. probably result from greater levels of assortative mating in this morph in comparison with the S-morph.
Similar content being viewed by others


Introduction
In the earlier paper of this pair (Baker et al., 2000), we described an unusual plant sexual polymorphism in which populations of Narcissus spp. (Amaryllidaceae), a genus of insect-pollinated geophytes native to the Mediterranean Basin, are dimorphic for style length. Most populations of N. assoanus and N. dubius in SW France consist of two floral morphs, one in which the stigma is at the same level as the stamens or protrudes beyond them (long-styled or L-morph) and the other in which the stigma is located well below the stamens (short-styled or S-morph). The polymorphism occurs in at least a dozen Narcissus species distributed across three sections of the genus (Barrett et al., 1996). Stigma-height dimorphism is distinct from the well-known heterostylous condition distyly (Darwin, 1877; Barrett, 1992), because stamen levels in the two floral morphs are positioned similarly, rather than showing a reciprocal correspondence with the two stigma heights. In addition, unlike most heterostylous species, the two floral morphs in Narcissus spp. do not differ in pollen size or pollen production.
Although N. assoanus and N. dubius show similar patterns of sex-organ deployment within flowers, population surveys in SW France revealed that the two species differ in the frequencies of style-morphs that occur within populations (Baker et al., 2000). In N. assoanus style-morph ratios ranged from isoplethy (1L:1S) to L-biased, depending on population size. Large populations were always isoplethic, whereas smaller populations exhibited variable style-morph frequencies, particularly L-biased ratios. In contrast, populations of N. dubius were either strongly L-biased or monomorphic for the L-morph. Of the 67 populations of the two species that were surveyed none was S-biased or contained only this morph. Similar patterns of style-morph frequency variation have also been reported in other dimorphic species of Narcissus with L-biased populations and L-monomorphy most commonly occurring (Arroyo & Dafni, 1995; Barrett et al., 1996).
In this paper we consider the maintenance of stigma-height dimorphism in Narcissus and investigate likely selective mechanisms that might account for the unusual patterns of morph-frequency variation observed in N. assoanus and N. dubius. We begin by determining the compatibility status of the two species through controlled pollination and then examine the possibility that morph-specific differences in maternal fitness of the style morphs could potentially lead to biased morph ratios. We compare estimates of the female fertility of style-morphs in natural populations by examining patterns of fruit- and seed-set. Style-morph ratios can also be influenced by morph-specific differences in mating patterns and an extensive literature on this topic is available for heterostylous plants (e.g. Crosby, 1949; Bodmer, 1960; Ganders, 1975; Charlesworth & Charlesworth, 1979; Barrett et al., 1987, 1989). We measure outcrossing rates in natural populations to determine whether morph-specific differences in floral morphology within each species might be associated with different patterns of mating. Finally, we develop a mating model to investigate how variation in levels of assortative and disassortative mating between the style morphs might influence their frequency within populations. In particular, we were interested in exploring the possibility that differences in assortative mating between style morphs caused by their differing sex-organ positions could explain the L-biased morph ratios that are typical of many Narcissus populations.
Materials and methods
All experimental studies described below were conducted from 1996 to 1998 on populations of N. assoanus and N. dubius occurring within 50 km of Montpellier, SW France. See figs 4 and 5 in Baker et al. (2000) for the general location of populations.
Compatibility status
We transplanted plants of the L- and S-morph of each Narcissus species to a pollinator-free glasshouse at CNRS, Montpellier, for hand-pollination to determine their compatibility status. It was particularly important to determine whether the species are self-sterile, and, if so, whether stylar dimorphism is associated with an intramorph incompatibility system typical of distylous plants. In N. assoanus, we conducted crosses over two years using three populations (St-Mathieu-de-Tréviers in 1996, and Bancel and Ceyrac in 1997). In N. dubius, it was difficult to find a substantial number of plants of the S-morph, even in large dimorphic populations. Thus, in N. dubius we performed the crossing programme in only one population in 1998.
In both species, the treatments were: (i) intramorph pollination; (ii) intermorph pollination; (iii) self-pollination; and (iv) no hand-pollination to detect the capability for autonomous selfing. All cross-pollinations involved a single, randomly chosen pollen donor. In both style morphs, we removed anthers from flowers prior to dehiscence and examined stigmas using a 16× hand lens to ensure that no self pollen was present before hand-pollination. It was necessary to slit open part of the floral tube to pollinate the S-morph, so flowers of both morphs were opened in the same manner. We collected mature fruits 6–8 weeks after pollination, counted the number of plump, filled seeds and flake-like ovules, and estimated total ovule number of flowers.
We analysed seed-set data from experimental pollinations by analysis of covariance (ANCOVA) in JMP (SAS, 1994) using total ovule number as a covariate. Only capsules that produced seed were included in the analysis. In N. assoanus, we examined the effects of style morph, pollination type (self-, intermorph and intramorph), population, and their interactions. Three-way interaction terms were backwards eliminated from the analyses if they did not have a significant effect on the mean number of seeds set (i.e. P > 0.05; cf. Sokal & Rohlf, 1995). We performed contrasts to compare self vs. outcrossed and intramorph vs. intermorph seed-set; and logistic regression to examine the effects of the above variables on mean fruit-set. We employed a similar approach in analysing seed- and fruit-set data in N. dubius. We used ANCOVA to examine the effects of style morph, pollination type and their interaction on mean seed-set. We performed contrasts as above and logistic regression to analyse the mean proportion of fruit-set.
Female fertility in natural populations
We measured seed-set of tagged L- and S-styled plants in six dimorphic N. assoanus populations in 1996 [Bancel (Ban), Col Hortus (Col), Hortus Mountain (Hmt), Pont sur Brestalou (Psb), Rigalderie (Rig) and St-Bauzille-de-Montmel (StB)], and both seed- and fruit-set in four populations in 1997 [Col, Psb, StB and Aguzan (Agu)]. Seed-set was measured in N. dubius for 1996 and 1998. In 1996, we sampled three populations exhibiting stigma-height dimorphism [La Clause (Clau), Hortus Mountain (Hmt), and St-Bauzille-de-Montmel (StB)] and two populations monomorphic for the L-morph [La Baune (Bau) and Pont du Gard (PdG)]. In 1998, we measured both fruit- and seed-set in a subset of these populations: Clau, StB and PdG.
In N. assoanus, we analysed seed-set data for both years using a mixed-model ANCOVA with total ovule number as the covariate. Random effects included year sampled and population nested within year. Style morph was treated as a fixed effect. Interaction terms in the model included morph by year and morph by population (nested within year). We analysed fruit-set data for 1997 using logistic regression. The two-way model examined the effects of morph, population and their interaction on fruit-set. In N. dubius, we analysed seed-set over the two years using a nested design. Effects included sampling year, population nested within year, and morph nested within population and year. Total ovule number was used as a covariate. We analysed the 1998 fruit-set data using logistic regression and examined the effects of population and morph nested within population on proportion of fruit set.
Mating patterns
We performed horizontal starch gel electrophoresis on open-pollinated seeds from six populations of N. assoanus (Ban, Col, Hmt, Psb, Rig, StB) and three populations of N. dubius (Clau, Hmt, StB), following the methods of Shore & Barrett (1987) and Wendel & Weeden (1989). All populations were dimorphic for style length. Electrophoresis was also performed on seed families from two monomorphic N. dubius populations (Bau, PdG). We initially screened all populations for 18 enzyme systems: from these, six polymorphic enzyme systems (Adh, Gdh, Idh, Got, Pgi, Tpi) were reliably resolved in N. assoanus and two (Acp, Pgm) in N. dubius. We used Ritland’s MLTR program (Ritland, 1990) to estimate morph-specific outcrossing rates. Outcrossing rates (t̂) and their standard deviations (based on 1000 bootstrap valves) were estimated from approximately 20 seed families per morph (range 15–21) for a given population. For each seed family, eight individual seeds were genotyped.
Ritland’s MLTR program assumes that allele frequencies do not differ significantly between groups, implying that mating is random with respect to morph (i.e. no disassortative mating between morphs). However, this seems highly unlikely given the 1:1 morph ratios of some populations which imply significant disassortative mating (Baker et al., 2000). If this assumption is violated, there will be a downward bias in t̂ when a common pollen pool is used to estimate the outcrossing rate rather than a separate pollen pool. Because trial runs indicated that there was a significant difference in allele frequencies between the morphs at some loci in all populations, outcrossing rates were estimated using separate pollen pools (and see Eckert & Barrett, 1994).
Differences in outcrossing rate between the style morphs in dimorphic populations were assessed by pairwise comparisons of bootstrap estimates following the method of Eckert & Barrett (1994). With this approach, morphs were considered to differ significantly if 100(1 − αPC/2) percentage of the differences between paired bootstrap values were either all greater than zero or all less than zero (note that αPC represents the type-I error rate per contrast). Differences in t̂ between populations were evaluated in a similar manner: 1000 average estimates of outcrossing were calculated for each population by calculating the average of the L- and S-morph t̂ for a given bootstrap and two populations were considered to differ significantly if, as above, 100(1 − αPC/2) percentage of the differences between paired bootstraps were either all greater than zero or all less than zero.
A genetic mating model
We developed a computer model to calculate equilibrium genotype and phenotype frequencies given varying levels of morph-specific assortative and disassortative mating. The explicit inclusion of assortative mating distinguishes this model from earlier mating models of heterostylous populations (e.g. Charlesworth, 1979; Heuch, 1979). Table 1 is a matrix the elements of which are the expected proportions of progeny genotypes from all matings for each maternal genotype. In our model, we assumed that the genetic control of stigma-height dimorphism is a single diallelic locus with the L-morph homozygous recessive (ss) and the S-morph Ss or SS, as reported for Narcissus tazetta (Dulberger, 1964 and unpubl. data). In each morph, matings were either assortative (within-morph) or disassortative (between-morph); for example in Table 1, aS represents the proportion of matings between S-plants (assortative) and dS represents the proportion of matings in which a maternal S-plant is pollinated by pollen from a plant of the L-morph (disassortative). We do not explicitly incorporate the rate of self-fertilization into our model because selfing can be considered genetically equivalent to assortative mating (i.e. the genotypes of selfed progeny are the same as progeny resulting from assortative mating).
In the model, we assumed initial genotype frequencies of 1ss (long):1Ss (short). Previous studies have not found short-styled SS individuals in natural populations of species with a stigma-height dimorphism (see discussion in Baker et al., 2000). In addition, large, presumably equilibrium populations of N. assoanus have equal frequencies of the two style morphs so we were interested to examine how variable mating patterns might change morph frequencies from this equilibrium. Calculations were run in which mating varied from complete disassortative mating (S=aL=0) to relatively high assortative mating (aS/dS=aL/dL=2) and genotype and phenotype frequencies were followed. Each set of calculations was run until an equilibrium in genotype frequency was reached (i.e. the per generation change in progeny genotype frequency was less than 0.0001). Results of the calculations were plotted in three-dimensions to illustrate the relationship between morph-specific mating in the L- and S-morphs and genotype frequency at equilibrium.
Results
Compatibility relationships
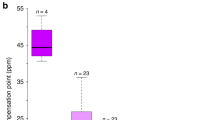
Controlled self- and cross-pollination indicates that N. assoanus is moderately self-sterile whereas N. dubius is highly self-compatible (Fig. 1). The similar levels of seed-set obtained following intra- and intermorph crosses in N. assoanus demonstrate that this species does not possess a heteromorphic incompatibility system.

Mean seed-set of the L- and S-morphs in three populations of Narcissus assoanus and one population of N. dubius following intra-, intermorph and self-pollination in the glasshouse. Narcissus assoanus exhibits moderate self-sterility, whereas N. dubius is self-compatible. There was no significant difference in seed-set between inter- and intramorph pollination in either species.
In N. assoanus, self-pollinations resulted in significantly fewer seed than intra- and intermorph cross-pollination in each of the three populations examined (grand mean seed-set: self=8.32, SE=0.82, n=57; intramorph=21.36, SE=1.32, n=74; intermorph= 18.18, SE=1.32, n=74). The results of paired contrasts indicated a significant difference in seed-set between self vs. outcross pollinations but not between the seed-set of intramorph vs. intermorph crosses (Table 2). Fruit-set in N. assoanus showed a similar pattern to the results obtained for seed-set. Mean fruit-set differed significantly between pollination treatments (χ22=16.89, P < 0.001) with higher fruit-set in outcrossed pollination (mean=77.9%, SE=4.3, n=95 for both intra- and intermorph pollination) compared with self-pollination (mean=55.9%, SE=4.9, n=102). Plants of neither morph set seed autonomously (n=8 for each morph in the St-Mathieu-de-Tréviers population) and this treatment was not included in the ANCOVA. An interesting feature of the compatibility data in N. assoanus is the significant effect of style morph on the number of seeds produced (Fig. 1; Table 2). Long-styled plants had significantly lower seed-set than S-plants and this pattern was consistent across all populations examined. Fruit-set was also reduced in the L-morph (mean=57.9%, SE= 4.0, n=152) when compared to the S-morph (mean= 83.6%, SE=3.1, n=140; χ21=17.51, P < 0.001).
There were no significant differences in seed-set between self-, intramorph and intermorph pollination of N. dubius (self=14.29, SE=1.48, n=38; intramorph=18.37, SE=1.74, n=38; intermorph=17.07, SE=1.38, n=41; Fig. 1, Table 3) and mean fruit-set was consistently high across all pollination types (self=84.4%, SE=5.5, n=45; intermorph=93.2%, SE=3.8, n=44; intramorph=88.4%, SE=4.9, n=43). There were no significant effects of pollination type, morph, or their interaction on fruit-set (χ25=3.64, P=0.6022). Unlike N. assoanus, there was no significant difference in seed-set between the style morphs in response to the pollination treatments (Fig. 1, Table 2). In spite of the lack of self-incompatibility, plants of N. dubius do not set fruit autonomously (n=17 and 4, respectively, for L- and S-styled plants).
Natural fertility
Comparisons of fruit- and seed-set of the L- and S-morphs in populations of N. assoanus and N. dubius failed to detect any general morph-specific effects. In only one of the 18 paired comparisons (N. dubius-StB-1996) was there a significant difference in the fertility of the style morphs (Fig. 2). In N. assoanus, mean seed-set varied significantly between years (1996=23.38, SE=0.65, n=458; 1997=13.92, SE=1.17, n=184; F1,555=34.62, P=0.0138) but style morph and population (nested within year) had no effect on mean seed-set (P > 0.13 in both cases). Fruit-set data were only available for populations sampled in 1997. Analysis of these data indicated no effect of style morph, population, or their interaction (χ27=9.09, P=0.2463).

Natural seed-set of the L- and S-morph in dimorphic populations of Narcissus assoanus and dimorphic and monomorphic populations of N. dubius over two flowering seasons. An asterisk indicates the population in which a morph-specific difference in seed-set was detected following contrasts (see text for details). Error bars indicate the standard error of the mean.
In N. dubius populations, style morph (nested within population and year) had a significant influence on seed-set (F5,310=3.25, P=0.0071). This effect was largely caused by the results from population StB in 1996 (L-morph=27.78, SE=1.96, n=35; S-morph=16.52, SE=4.32, n=7; contrast t5=3.93, P=0.0001). However, when we performed contrasts to compare seed-set between the style morphs in each dimorphic population, no other significant differences were evident (Clau in 1996 and 1998, Hmt in 1996, and StB in 1998; P > 0.56 for all contrasts). Population nested within year also had a significant effect on mean seed-set (F6,310=3.03, P=0.0067), although there was no significant difference in mean seed-set between years (F1,310=0.03, P=0.8659). Fruit-set data were only available for populations sampled in 1998. There was no evidence for morph-specific differences in mean fruit-set (L-morph=87.02%, SE=3.00, n=75; S-morph= 79.29, SE=6.71%, n=19) or of differences in fruit-set between populations (χ24=6.25, P=0.1809).
Mating patterns of style morphs in natural populations
Comparisons of outcrossing rate in N. assoanus and N. dubius populations failed to detect significant differences between the L- and S-morphs (Fig. 3). In no case were all pairwise comparisons of morph-specific bootstrap values above or below zero. Populations of N. assoanus and N. dubius exhibited contrasting mating systems. Narcissus assoanus is highly outcrossing (mean t̂: L-morph=0.997, SD=0.0038; S-morph=0.999, SD=0.0014), whereas N. dubius exhibits a mixed mating system (mean t̂ in dimorphic populations: L-morph= 0.579, SD=0.071; S-morph =0.653, SD=0.099). The larger variances in t̂ values obtained for populations of N. dubius compared with N. assoanus (Fig. 3) undoubtedly result, in part, from the small number of allozyme loci available for these estimates (two loci in N. dubius and six loci in N. assoanus). Comparisons of t̂ among N. assoanus populations indicated no significant differences. Only one of the pairwise comparisons among N. dubius populations was significant: t̂ in the monomorphic PdG population was significantly lower than in the dimorphic Hmt population (PDG mean t̂=0396, SD=0.085; Hmt mean t̂=0.735, SD=0.077).

Estimated multilocus outcrossing rates (t̂) of the L- and S-morph in populations of Narcissus assoanus and N. dubius. Morph-specific estimates were obtained using separate pollen pools in Ritland’s (1990) MLTR program and error bars are standard deviations based on 1000 bootstrap estimates. Within each dimorphic population there were no morph-specific differences in mating system in either species.
Effects of assortative and disassortative mating on style-morph frequencies
The results of computer calculations on the effects of assortative and disassortative mating on style-morph frequencies are illustrated in Fig. 4. With complete disassortative mating (aS/dS=aL/dL=0) equilibrium morph ratios in a population were 1L:1S (Fig. 4a,b) and all individuals of the S-morph were heterozygous at the locus controlling style length. Stylar dimorphism was maintained when disassortative mating occurred more frequently than assortative mating and the rate of disassortative mating was equivalent among the morphs (aL/dL and aS/dS < 1; Fig. 4a). When these conditions were met, morph ratios did not differ from 1L:1S and the genotype of the S-morph was primarily Ss (Fig. 4b), although the frequency of the SS genotype increased as assortative mating in plants of the S-morph (aS) increased (Fig. 4c).

Equilibrium genotype frequencies with varying amounts of assortative and disassortative mating in the style morphs: (a) frequency of L-morph (ss); (b) frequency of the S-morph (Ss); and (c) frequency of the S-morph (SS). aL/dL and aS/dS represent the ratio of assortative to disassortative mating in the maternal parent such that aL/dL and aS/dS > 1 is high assortative mating in both morphs and aL/dL=aS/dS=0 is complete disassortative mating. Note that the orientation of the x- (aL/dL) and y- (aS/dS) axes differs between graphs for optimal viewing of the surface representing genotype frequency. See text for further details of the model.
The computer calculations indicated when L-biased morph ratios would occur. When assortative mating was low in the S-morph (aS/dS < 0.8) but intermediate in frequency in the L-morph (aL/dL=0.8–1.2), equilibrium morph ratios were biased towards the L-morph and populations became fixed for this morph if the proportion of assortative matings in the L-morph increased (Fig. 4a). At very high levels of aS or aL, populations became fixed for the S- or L-morph, respectively. This is because assortative mating in the S-morph results in the segregation of the SS genotype and assortative mating in the L-morph results only in L-styled offspring. S-monomorphy occurred when assortative mating was high in the S-morph (aS/dS > 1.7) but low in the L-morph (Fig. 4a). Under this particular scenario, the genotypes of S-individuals were primarily SS (Fig. 4b, c).
Discussion
This study provides additional insight into the nature and maintenance of stigma-height dimorphism in Narcissus. The polymorphism in N. assoanus and N. dubius differs from distyly because neither species possesses reciprocal herkogamy, pollen heteromorphisms or heteromorphic incompatibility. These facts support the argument that the dimorphism is fundamentally distinct from heterostyly (Baker et al., 2000). Yet the stigma-height dimorphism is manifested quite differently in the two Narcissus species because of features of their floral biology that influence the mating systems of populations. Here we discuss how contrasting physiological and morphological traits in N. assoanus and N. dubius affect mating and fertility and result in the patterns of morph-frequency variation observed in populations of the two species in SW France.
Compatibility status of Narcissus assoanus and N. dubius
Our controlled glasshouse pollinations indicated that whereas N. assoanus is moderately self-sterile, N. dubius is thoroughly self-compatible. Most Narcissus species that have been examined experimentally have proved to be self-incompatible (Bateman, 1954; Dulberger, 1964; Barrett et al., 1996, 1997; Sage et al., 1999), although several fully self-compatible species are reported (Herrera, 1995; Barrett et al., 1996). Despite the presence of stylar polymorphisms in Narcissus there is no evidence that any species in the genus possesses heteromorphic incompatibility. However, the class of incompatibility (gametophytic vs. sporophytic) and its genetic basis (single-locus vs. multilocus) remain unclear. Studies of dimorphic N. tazetta and trimorphic N. triandrus indicate a late-acting self-incompatibility system in which self and cross pollen tubes grow at equivalent rates and self-rejection occurs in the ovary (Bateman, 1954; Dulberger, 1964; Barrett et al., 1997; Sage et al., 1999). It is possible that N. assoanus also possesses late-acting incompatibility because, in common with these species, preliminary crosses have indicated that prior self-pollination of flowers 48 h before outcrossing results in a significant reduction in fertility (A.M. Baker, J.D. Thompson & S.C.H. Barrett, unpubl. data). However, the reduced seed-set upon selfing could also arise from early acting inbreeding depression (Seavey & Bawa, 1986; Husband & Schemske, 1996). Detailed investigations of pollen–pistil interactions and ovule and seed development would be required to confirm this (e.g. see Sage et al., 1999).
All three populations of N. assoanus responded similarly to the pollination treatments. There was considerable variation among plants in their response to self-pollination, with some strongly self-sterile and others in which self-sterility was only partially expressed. A few individuals were detected that set equivalent amounts of seed from both self- and cross-pollinations. There was no evidence that the style morphs differed in the overall strength of self-sterility that might influence selfing rates. Data on mating patterns in natural populations were consistent with this finding because no differences in outcrossing rate between style morphs were observed (see below). In N. dubius, most individuals set abundant seed on self-pollination. Self-compatibility in this species may have arisen through selection of individuals in ancestral populations with weak expression of self-sterility similar to that observed in N. assoanus. Uncertain pollinator service could explain the self-compatible status of this early spring-flowering species. Narcissus longispathus, another early flowering species, is also highly self-compatible (Herrera, 1995), and early flowering has often been invoked to explain the evolution of increased levels of self-fertilization (Lloyd, 1965; Jain, 1976).
An unexpected feature of the crossing data for N. assoanus was the significantly lower fertility of the L-morph in comparison with the S-morph (Fig. 1). On average, plants of the L-morph set 33% less seed and 31% fewer fruit from cross-pollinations than those of the S-morph. This pattern was evident in all three populations used in the crossing programme (data not shown) in two separate years. Yet our measurements of the natural fertility of six N. assoanus populations, including one of the three populations used in glasshouse pollinations, failed to detect significant morph-specific differences in female fertility. It seems unlikely that this result is caused by sampling effects associated with the particular plants of the L-morph used in the glasshouse studies as sample sizes were generally large. Also, the contrasting patterns of fertility cannot result from ovule discounting because of the influence of different degrees of herkogamy on autonomous self-pollination (see Barrett et al., 1996; Baker et al., 2000). All flowers used in hand-pollinations were emasculated prior to being cross-pollinated. It is possible that environmental conditions in the glasshouse influenced the fertility of the style morphs differently; for example, if temperature and humidity conditions were suboptimal for pollen–pistil interactions, fewer pollen tubes may have reached receptive ovules in the L-morph in comparison with the S-morph because of differences between the morphs in style length. Of relevance to this interpretation is the finding that the time taken for pollen tubes to reach ovules is longest in the L-morph and shortest in the S-morph in tristylous N. triandrus (Sage et al., 1999).
Maternal fitness components in natural populations
Style-morph ratios in populations of Narcissus species are likely to be more sensitive to morph-specific differences in fitness compared with species in which disassortative mating is guaranteed because of heteromorphic incompatibility. We therefore sought to establish whether differences in maternal fitness components of the style morphs could explain the wide ranges of population morph frequencies that occur in N. assoanus and N. dubius. However, our measurements of female fertility and outcrossing rate in natural populations failed to detect differences between the style morphs. Thus it is unlikely that these features of the reproductive biology are of any significance in accounting for the patterns of morph-ratio variation previously documented (Baker et al., 2000). Although occasional differences between morphs within particular populations were sometimes detected, there was no evidence of any clear trends that could account for the L-biased ratios that commonly occur.
Within populations of N. assoanus levels of fruit- and seed-set of the style morphs were similar, despite striking variation among years. Outcrossing rates of the morphs were near 1.0 in all populations, indicating that despite the potential for high levels of autonomous self-pollination in the L-morph; self-sterility apparently prevents both increased ovule discounting and selfing in this morph. How this is achieved is by no means clear. Previous studies demonstrated that prior selfing can reduce fertility in this and other Narcissus species (Dulberger, 1964; Barrett et al., 1997; A.M. Baker, J.D. Thompson & S.C.H. Barrett, unpubl. data), leading us to expect that autonomous self-pollination might result in reduced female fertility in the L-morph in comparison with the S-morph. This could occur if autonomous self-pollination renders ovules unreceptive prior to the arrival of outcrossed pollen. This expectation was not borne out, suggesting that either sufficient outcross pollen is delivered to L-stigmas by pollinators before autodeposition occurs, or that postpollination mechanisms act to favour outcross fertilizations and hence maintain fertility.
Although our sampling of N. dubius was more limited, there were no consistent fertility differences between the style morphs that seem likely to account for the strongly L-biased morph ratios observed in this species. In one population (StB) in 1996, the L-morph set 68% more seed than the S-morph. If this difference were to occur regularly, it could influence the dynamics of morph frequencies in this population. However, in 1998, comparisons of the fertility of style morphs in this population failed to detect the pattern observed in 1996, so it is not clear how often such an effect occurs. Estimates of outcrossing in the three dimorphic populations that were examined also failed to reveal significant style morph effects, although the power to detect small differences in mating was low because of the limited number of allozyme loci (two) available for the estimates. Overall, N. dubius exhibited a mixed-mating system with an average of 42% of seed arising from self-fertilization. Higher selfing in N. dubius, compared to N. assoanus, was not unexpected given the self-compatible status and the possession of multiflowered inflorescences. Even the relatively small floral displays found in N. dubius (one to seven flowers per inflorescence) provide opportunities for geitonogamy that are not possible in most N. assoanus plants, which are largely solitary-flowered in SW France. The occurrence of several inflorescences per plant in N. dubius because of bulb fragmentation also increases the likelihood of geitonogamous selfing.
Pollen dispersal and its influence on mating patterns
Our failure to detect differences in maternal fitness components between the style morphs of N. assoanus and N. dubius motivated us to consider the influence of pollen dispersal on mating patterns. Unfortunately, measurement of the key parameters required to understand the relationships between floral morphology, pollen dispersal and mating was hampered by the absence of pollen-size heteromorphism in Narcissus (Baker et al., 2000), and a lack of suitable genetic markers for estimating pollen transfer and levels of assortative and disassortative mating. Because of these difficulties, we chose to adopt a theoretical approach by developing a simple genetic model that explored the relationships between mating patterns and style-morph ratios. The model demonstrated that stigma-height dimorphism will always be maintained when levels of disassortative mating are greater than assortative mating, and that morph-specific differences in these mating parameters give rise to populations with biased style-morph ratios. These findings imply that the commonly observed L-biased morph ratios in Narcissus spp. result from higher levels of assortative mating in the L- vs. the S-morph. How likely is this to occur?
Several lines of evidence support the view that assortative mating will be more frequent in the L-morph in comparison to the S-morph. Because most monomorphic species of Narcissus have sex organs in a similar position to the L-morph it seems reasonable to assume that effective pollen transfer between plants with this morphology is generally adaptive. The common occurrence of populations monomorphic for the L-morph, but not the S-morph, among dimorphic Narcissus species is consistent with this view (Barrett et al., 1996; Baker et al., 2000). Thus, in dimorphic populations, matings between plants of the L-morph should be common. However, although stigmas of the L-morph correspond in height to upper-level anthers of both morphs, stigmas of the S-morph are situated well below the lower-level anthers, particularly those of the S-morph (Baker et al., 2000). The disparity in organ height may reduce the precision of intramorph pollinations in the S-morph. Indeed, experimental studies of heterostylous plants have demonstrated that pollen transfer between anthers and stigmas of equivalent height occurs much more often than between sex organs of dissimilar position (Ganders, 1974; Barrett & Glover, 1985; Kohn & Barrett, 1992; Lloyd & Webb, 1992). Therefore it seems likely that the high degree of herkogamy in the S-morph, compared to the L-morph, reduces the likelihood of assortative mating in this morph. According to this interpretation, most matings in the S-morph are disassortative, whereas a mixture of assortative and disassortative mating is likely to occur in the L-morph. As our simulations demonstrate, this mating pattern results in populations with L-biased morph ratios or populations fixed for the L-morph. This mating pattern also predicts that plants of the S-morph will largely be of the Ss genotype, a situation consistent with genetic data obtained from N. tazetta (Dulberger, 1964; see Baker et al., 2000).
In conclusion, populations of N. assoanus and N. dubius exhibit a continuum of style-morph frequencies from isoplethy (1:1) to populations fixed for the L-morph. Narcissus assoanus populations are always dimorphic and some occur at the isoplethic end of the continuum, whereas populations of N. dubius are strongly L-biased and occasionally fixed for this morph. We interpret this variation as resulting from differences between the two species in mating patterns, particularly levels of assortative and disassortative mating. Self-sterility in N. assoanus favours the maintenance of stigma-height dimorphism because the vast majority of matings arise from outcrossing, and in many populations, a sizeable fraction must arise from crosses between the style morphs. In contrast, in N. dubius, disassortative mating appears to be more difficult to achieve and self-compatibility may have aided secondary reversion to stylar monomorphism in some populations through founder events. These contrasting influences on style-morph ratios in Narcissus spp. serve to illustrate the complex dynamics that can occur when morphological and physiological traits are not genetically associated as in most heterostylous species. Because of this, stigma-height polymorphisms provide excellent opportunities to investigate natural selection on simply inherited floral traits without the confounding effects of heteromorphic incompatibility.
References
Arroyo, J. and Dafni, A. (1995). Variation in habitat, season, flower traits, and pollinators in dimorphic Narcissus tazetta L. (Amaryllidaceae) in Israel. New Phytol, 129: 135–145.
Baker, A. M., Thompson, J. D. and Barrett, S. C. H. (2000). Evolution and maintenance of stigma-height dimorphism in Narcissus I. Floral variation and style-morph ratios. Heredity. 84: in press.
Barrett, S. C. H. (ed.) (1992). Evolution and Function of Heterostyly. Springer-Verlag, Berlin.
Barrett, S. C. H. and Glover, D. E. (1985). On the Darwinian hypothesis of the adaptive significance of tristyly. Evolution 39: 766–774.
Barrett, S. C. H., Brown, A. H. D. and Shore, J. S. (1987). Disassortative mating in tristylous Eichhornia paniculata (Spreng.) Solms (Pontederiaceae). Heredity 58: 49–55.
Barrett, S. C. H., Morgan, M. T. and Husband, B. C. (1989). The dissolution of a complex genetic polymorphism: the evolution of self-fertilization in tristylous Eichhornia paniculata (Pontederiaceae). Evolution 43: 1398–1416.
Barrett, S. C. H., Lloyd, D. G. and Arroyo, J. (1996). Stylar polymorphisms and the evolution of heterostyly in Narcissus (Amaryllidaceae). In: Lloyd, D. G. and Barrett, S. C. H. (eds) Floral Biology. Studies on Floral Evolution in Animal-Pollinated Plants pp. 339–376. Chapman & Hall, New York.
Barrett, S. C. H., Cole, W. W., Arroyo, J., Cruzan, M. B. and Lloyd, D. G. (1997). Sexual polymorphisms in Narcissus triandrus (Amaryllidaceae): is this species tristylous? Heredity 78: 135–145.
Bateman, A. J. (1954). The genetics of Narcissus I. Sterility. In: Daffodil and Tulip Yearbook 19 pp. 23–29. Royal Horticulture Society, London.
Bodmer, W. F. (1960). Genetics of homostyly in populations of Primula vulgaris. Phil Trans R Soc B, 242: 517–549.
Charlesworth, D. (1979). The evolution and breakdown of tristyly. Evolution 33: 486–498.
Charlesworth, D. and Charlesworth, B. (1979). A model for the evolution of distyly. Am Nat, 114: 467–498.
Crosby, J. L. (1949). Selection of an unfavourable gene complex. Evolution, 3: 212–230.
Darwin, C. (1877). The Different Forms of Flowers on Plants of the Same Species Reprinted in 1986. University of Chicago Press, Chicago, IL.
Dulberger, R. (1964). Floral dimorphism and self-incompatibility in Narcissus tazetta L. Evolution, 18: 361–363.
Eckert, C. G. and Barrett, S. C. H. (1994). Post-pollination mechanisms and the maintenance of outcrossing in self-compatible, tristylous, Decodon verticillatus (Lythraceae). Heredity, 72: 396–411.
Ganders, F. R. (1974). Disassortative pollination in the distylous plant Jepsonia heterandra. Can J Bot, 52: 2401–2406.
Ganders, F. R. (1975). Mating patterns in self-compatible distylous populations of Amsinckia (Boraginaceae). Can J Bot, 53: 773–779.
Herrera, C. M. (1995). Floral biology, microclimate, and pollination by ectothermic bees in an early-blooming herb. Ecology, 76: 218–228.
Heuch, I. (1979). Equilibrium populations of heterostylous plants. Theor Pop Biol, 15: 43–57.
Husband, B. C. and Schemske, D. W. (1996). Evolution of the magnitude and timing of inbreeding depression in plants. Evolution 50: 54–70.
Jain, S. K. (1976). The evolution of inbreeding in plants. Ann Rev Ecol Syst, 7: 69–95.
Kohn, J. R. and Barrett, S. C. H. (1992). Experimental studies on the functional significance of heterostyly. Evolution, 46: 43–55.
Lloyd, D. G. (1965). Evolution of self-compatibility and racial differentiation in Leavenworthia (Cruciferae). Contr Gray Herb, 195: 3–133.
Lloyd, D. G. and Webb, C. J. (1992). The selection of heterostyly. In: Barrett, S. C. H. (ed.) Evolution and Function of Heterostyly pp. 179–207. Springer-Verlag, Berlin.
Ritland, K. (1990). A series of Fortran computer programs for estimating plant mating systems. J Hered, 81: 235–237.
Sage, T. L., Strumas, F., Cole, W. W. and Barrett, S. C. H. (1999). Differential ovule development following self- and cross-pollination: the basis of self-sterility in Narcissus triandrus (Amaryllidaceae). Am J Bot, 86: 855–870.
SAS (1994). JMP Users Guide version 3.0.2. SAS Institute, Cary, NC.
Seavey, S. R. and Bawa, K. S. (1986). Late-acting self-incompatibility in angiosperms. Bot Rev, 52: 195–219.
Shore, J. S. and Barrett, S. C. H. (1987). Inheritance of floral and isozyme polymorphisms in Turnera ulmifolia L. J Hered 78: 44–48.
Sokal, R. R. and Rohlf, F. J. (1995). Biometry 3rd edn. W. H. Freeman & Co., San Francisco, CA.
Wendel, J. F. and Weeden, N. F. (1989). Visualization and interpretation of plant isozymes. In: Soltis, D. E. and Soltis, P. S. (eds) Isozymes in Plant Biology pp. 5–45. Dioscorides Press, Portland, OR.
Acknowledgements
We thank Lawrence Harder and Anne Worley for valuable discussion and assistance in the field; Peter Taylor, Chris Eckert, Linley Jesson and Bill Cole for advice in the development of the mating model and comments on the manuscript; and Stephen Wright for assistance in seed counting. This work was funded by an operating grant from the Natural Sciences and Engineering Research Council of Canada (NSERC) to S.C.H.B. and a grant from CNRS to J.D.T. A.M.B. was supported by graduate scholarships from NSERC and the Ontario government.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Baker, A., Thompson, J. & Barrett, S. Evolution and maintenance of stigma-height dimorphism in Narcissus. II. Fitness comparisons between style morphs. Heredity 84, 514–524 (2000). https://doi.org/10.1046/j.1365-2540.2000.00686.x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1046/j.1365-2540.2000.00686.x
