
Abstract
An understanding of mating systems and fine-scale spatial genetic structure is required to effectively manage forest pest species such as Dendroctonus ponderosae (mountain pine beetle). Here we used genome-wide single-nucleotide polymorphisms to assess the fine-scale genetic structure and mating system of D. ponderosae collected from a single stand in Alberta, Canada. Fine-scale spatial genetic structure was absent within the stand and the majority of genetic variation was best explained at the individual level. Relatedness estimates support previous reports of pre-emergence mating. Parentage assignment tests indicate that a polygamous mating system better explains the relationships among individuals within a gallery than the previously reported female monogamous/male polygynous system. Furthermore, there is some evidence to suggest that females may exploit the galleries of other females, at least under epidemic conditions. Our results suggest that current management models are likely to be effective across large geographic areas based on the absence of fine-scale genetic structure.
Similar content being viewed by others


Introduction
The mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera: Curculionidae), is a native forest pest throughout western North America (Maness et al., 2012). Its pre-2005 range extended from northern Mexico to central British Columbia and southwestern Alberta, and an isolated population was located in the Cypress Hills in southeastern Alberta and Saskatchewan, Canada (Mock et al., 2007; Safranyik et al., 2010). Dendroctonus ponderosae is one of the few North American bark beetles that can kill live, healthy trees (primarily Pinus species) in order to feed and complete its life cycle (Lee et al., 2006; Reeve et al., 2012). Populations naturally cycle between phases of endemic (small, low-density populations attacking low-quality hosts) and epidemic (wide-spread, high-density populations attacking mature healthy trees) (Safranyik and Wilson, 2006) approximately every 20 years (Aukema et al., 2006; Safranyik et al., 2010). Typically, these epidemic phases have been regulated by natural predators (for example, woodpeckers; Fayt et al., 2005), interspecific competition (for example, Ips pini; Rankin, 1988), intraspecific competition (Trzcinski and Reid, 2009) and temperature (Régnière and Bentz, 2007). However, in recent years warmer temperatures and altered forest management practices have resulted in what is possibly the biggest epidemic outbreak recorded in history (Taylor and Carroll, 2004).
Around 2005, D. ponderosae began to invade boreal Canada and spread throughout much of north-central Alberta and into the southern Northwest Territories (Lee et al., 2006; Safranyik et al., 2010). This outbreak has devastated over 18 million hectares of forest (Aukema et al., 2008; Natural Resources Canada, 2015a), causing significant economic, social and environmental damage (Aukema et al., 2006). The range expansion raised concerns that D. ponderosae would continue to spread through the boreal. A hybrid zone of lodgepole pine × jack pine in parts of north central Alberta (Cullingham et al., 2011) provides a potential bridge, allowing D. ponderosae to become better suited to jack pine as a host, thereby providing access to the boreal forests across North America. Resultantly, management of D. ponderosae in boreal forests is of national importance (Natural Resources Canada, 2015b); however, initial research has shown that the biology (that is, brood development and survival) of D. ponderosae populations in the boreal forest is different to populations in western and southern forest types (Rice et al., 2008; Safranyik et al., 2010; Myrholm and Langor, 2015). These differences highlighted the potential risk of simply extending management based on historical knowledge of D. ponderosae directly to the boreal. It was clear that effective management of D. ponderosae in the boreal required new research about mountain pine beetle biology and its ecological interactions within boreal systems (Safranyik et al., 2010), including information about life cycle, mortality, dispersal and mating systems.
Throughout much of its historical range in Canada, female beetles typically initiate host attack and egg gallery construction between July and August (Safranyik and Wilson, 2006). Males join females later, attracted by female-emitted pheromones, and mating occurs in the galleries (Safranyik and Wilson, 2006). Larvae develop and pass through four instars, overwintering largely in the third and fourth instar, before pupating in June (Aukema et al., 2006). Teneral adults undergo hardening before emerging from the host tree gallery and embarking on their own dispersal flights in June and July of the following year (Aukema et al., 2006). However, observations suggest that these events occur earlier in boreal systems as less thermal units are required to achieve a generation (Bentz et al., 2014; DW Langor, unpublished data). Dendroctonus ponderosae populations typically exhibit univoltine reproduction (Sturgeon and Mitton, 1986).
The mating system of D. ponderosae is complex. Males join females in freshly constructed egg galleries where mating takes place (Reid, 1962). However, recent reports suggest that only 1–2% (Safranyik and Wilson, 2006) or up to 3–12% (Bleiker et al., 2013) of females have mated before they emerge from maternal galleries (that is, pre-emergence mating). It has been widely accepted that male beetles may be polygynous but there is no clear published indication that females engage in polyandry. Safranyik and Wilson (2006) state that sperm remains viable in the spermatheca for at least a year, suggesting that females would not have a biological need to mate again even if they survived winter and continued reproducing in the following year. In order to contribute to D. ponderosae management at the front of the boreal invasion in Alberta, and build on models predicting patterns and rates of invasion and success of D. ponderosae populations, a detailed understanding of beetle mating systems and resultant fine-scale population genetic structure would be extremely useful.
Studies of parentage and sibship relationships within D. ponderosae populations appear to be absent in the literature, presumably because of difficulties in conducting field-based studies (Berger-Wolf et al., 2007). However, information on genealogical relationships can be particularly valuable in studies of mating systems and spatial genetic structure in natural populations (Wang and Santure, 2009). The level of kinship among individuals within a population can be used to infer the predominant mating system (Berger-Wolf et al., 2007) and show the local pedigree structure that, in turn, provides information about the level of spatial genetic structure present (Jacquemyn et al., 2006). With the rapid development and growing application of molecular markers, studies into these areas are far more tractable. Single-nucleotide polymorphisms (SNPs) are particularly informative. Here we investigate the fine-scale genetic structure and mating system of D. ponderosae using SNPs. More specifically, we aim to determine whether D. ponderosae shows genetic evidence of: (1) fine-scale population genetic structure, (2) mating systems other than polygyny (that is, are females also polyandrous?) that may influence dispersal patterns and fine-scale population genetic structure and (3) pre-emergence mating.
Materials and methods
Specimen collection and DNA extraction
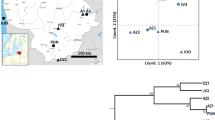
Adult beetles and larvae were collected from a boreal stand of lodgepole pine (Pinus contorta Douglas ex Loudon) trees near Fairview, Alberta (56.490 N, −118.518 W). Sampled trees were attacked by outbreak populations of D. ponderosae in July and August 2008 and were felled on 3 September 2008. Five trees ranging in height from 13.5 to 21.4 m were felled. Each tree was delimbed and cut to provide five sections with the following designations and heights above ground level: ST=0.5–1.0 m; B=2.0–3.0 m; D=4.0–5.0 m; F=6.0–7.0 m; and H=8.0–9.0 m. Sections were transported back to the laboratory where the bark was removed to expose individual egg gallery systems. These samples were given unique identifiers that would allow them to be traced back to individual trees, sections and galleries (that is, tree no.–section no.–gallery no.). Separately, to test for relatedness among proximate sites (within 750 km), additional adult beetles were genotyped: 11 beetles from Fort St James, British Columbia (56.704 N, −121.712 W; collected 2005); 20 from Grande Prairie, Alberta (54.992 N, −118.614 W; collected 2007); and 20 from Fox Creek, Alberta (54.481 N, −116.635 W; collected 2007) (Figure 1). Adults and larvae were rinsed in sterile distilled water and stored at −20 °C.

Map of beetle sites used in this study. The inset box shows the spatial distribution of the five trees sampled at Fairview.
Excavated galleries often contained adults, larvae or both. Where possible, gallery systems containing one or more adult beetles and late-instar larval progeny were selected for DNA extraction to best include potential parents. Thus, we used a number of galleries in which both adults and larvae were present to assess the mating system through parentage assignment; a number of galleries that contained only adults or larvae were then added to the analysis to further test the assignment and relatedness. The sex of adults was determined using characters of posterior margin of the seventh tergite (Safranyik and Wilson, 2006). Larvae could not be sexed because of an absence of diagnostic characteristics. Genomic DNA was extracted from adult and larval D. ponderosae specimens using QIAGEN (Toronto, ON, Canada) DNeasy 96 Blood & Tissue kits as per the manufacturer’s instructions, with modification to the lysis step. Lysis was completed by incubating the samples at 56 °C overnight. All DNA samples were checked for quality and normalized using Qubit (Invitrogen, Life Technologies, Carlsbad, CA, USA) flourometry and milliQ to a concentration of 20 ng μl−1 before genotyping.
Genotyping and filtering
A panel of 1536 SNPs was previously developed using the Illumina (San Diego, CA, USA) GoldenGate assay (Janes et al., 2014). From this GoldenGate panel, we selected 114 SNPs to develop a Sequenom panel. In total, 404 samples from the five Fairview trees (see Supplementary Table 1) were genotyped at McGill University and Génome Québec Innovation Centre using the Sequenom iPLEX Gold genotyping assay (Sequenom, 2008).
Genotyped samples and SNPs were filtered to ensure high quality and reproducibility. First, the quality of SNP and sample calls was assessed using the call rate. SNPs and samples with a call rate of ⩾80% were considered ‘good’, whereas SNPs and samples with call rates <80% were removed. Second, samples were checked for reproducibility. Third, samples and SNPs with >10% missing information (that is, NN) were removed.
Data analysis
Observed (HO) and expected (HE) heterozygosities were assessed using GenePop Version 4.1.3 (Rousset, 2008) to detect deviations from Hardy–Weinberg equilibrium. A Bonferroni adjustment (Holm, 1979), using an initial α of 0.05, was used to assess the statistical significance of multiple P-values. Outlier detection was used to identify SNPs under directional selection. BayeScan was run using the methodology described in Janes et al. (2014). To investigate fine-scale genetic structure and variation, the total genetic variance was assessed using a hierarchical analysis of molecular variance in GenAlEx 6.4 (Peakall and Smouse, 2006). Data were organized according to the following hierarchical partitions: (1) within individuals, among individuals within galleries (Fis) and among galleries (Fst); (2) within individuals, among individuals within sections from individual trees (Fis) and among sections within individual trees (Fst); (3) within individuals, among individuals within trees (Fis) and among trees (Fst); (4) within individuals (Fis), among individuals within galleries (Fst), among galleries within sections (Fsr) and among sections (Frt); and (5) within individuals (Fis), among individuals within sections (Fst), among sections within trees (Fsr) and among trees (Frt). Tests for significant departure from the null hypothesis that subpopulations are part of a single large, random mating, genetic population were performed using 999 random permutations.
Pairwise relatedness estimates were calculated using SPAGeDi 1.4 (Hardy and Vekemans, 2002). These relatedness estimates assess the biological relationships among individuals based on genotypic similarity through the proportion of shared alleles by calculating allele frequencies across the entire population, and standardizing average relatedness to a mean of zero. Therefore, a positive value between two individuals implies they are more related than expected by chance, whereas a negative value implies that they share fewer alleles than expected relative to the total population. The Fairview population was compared with the following spatially proximal populations: Grande Prairie, Fort St James and Fox Creek. To prevent biases from uneven sampling, allele frequencies from GenAlEx were averaged and used as an input for SPAGeDi.
Parentage and sibship were assigned using CERVUS 3.0.7 (Kalinowski et al., 2007) with the following parameters: (1) proportion of candidate mothers/fathers sampled 0.5; (2) proportion of loci mistyped 0.01; (3) strict confidence level 95%; and (4) accepted number of mismatches ⩽2. These assignments were performed for galleries containing putative parents with larvae, and for all individuals separately. Probability of identity was calculated using CERVUS.
Results
Fine-scale spatial genetic structure
In total, 2155 individuals were harvested, providing an average density of 431 (s.e.±0.133) and 86 (s.e.±0.062) individuals per tree and section, respectively. Because of limited availability of gallery systems containing one or more presumed parent adults with larvae, several galleries were selected that contained only larvae or adults. In total, 404 individuals (203 larvae and 201 adults) were selected for genotyping. Of the 201 adults, 140 were female, 56 were male and 5 were of undetermined sex. Adult females and males were found in galleries 80% and 32% of the time, respectively. These 404 individuals represented 174 galleries from 24 cross-sections across the 5 trees (one section did not yield suitable material). From these 174 galleries, 15 (8.6%) contained both adults and larvae, 15 (8.6%) did not have an adult present and 144 (82.7%) did not have larvae present. The 15 galleries that contained both adults and larvae yielded 119 larvae and 22 adults (11 female and 11 male), and were used to perform initial parentage assessments. Later, all 404 individuals were used to test for relatedness and further assess the mating system through parentage analyses. Using a call rate threshold of >80%, and removing SNPs and samples with >10% missing information, 21 (15.7%) SNPs were removed. The sample reproducibility error rate was <5%. Of the remaining 93 SNPs, 2 were monomorphic and removed, leaving a total of 91 SNPs for analyses. These 91 SNPs comprised 44 exonic, 25 intronic and 22 noncoding SNPs. Outlier detection tests revealed one directionally selected locus. The inclusion of this locus did not change the results obtained. We retained all 404 samples in our analyses.
Tests for deviation from Hardy–Weinberg proportions indicated that there were no significant deviations after sequential Bonferroni correction. All hierarchical analysis of molecular variance results (Table 1) suggest that the majority of molecular variance is best explained at the individual level, typically 76–78%. Very little genetic variance was explained at the individual tree (1%) or section levels (2–5%), with a modest degree explained at the level of galleries (10–12%). Mean pairwise Queller and Goodnight relatedness estimates within each population ranged from −0.04 to 0.01 and −0.021 to 0.001 among populations. Individuals within Fort St James were significantly less related to each other than expected when compared with individuals from within Fairview. Among populations, Fort St James and Grande Prairie were significantly less related to Fairview than the Fox Creek population. Figure 2 provides the estimates for each grouping with 95% confidence intervals.

Mean relatedness estimates (Queller and Goodnight) within and among populations showing 95% confidence limits. FC, Fox Creek; FSJ, Fort St James; FV, Fairview; GP, Grande Prairie.
Mating system
We identified six galleries from two trees that contained both males and females in the presence of larvae; an additional five galleries across four trees were found with females and larvae, and four galleries from two trees were found with males and larvae. One gallery (gallery 5-5-170; that is, tree no.—section no.—gallery no.) contained two males in the same gallery with larvae, but no female. These galleries contained a total of 119 larvae. In all, 6 of the 11 (54.55%) females and 8 of the 11 males (72.72%) were excluded from being the mother/father of at least one of the larvae found in their respective galleries. Three males (FV0993, FV1025 and FV1031) were found to be unrelated to all of the larvae found within their respective galleries. The gallery 5-5-166 contained both a male and female; however, the male (FV0993) was unrelated to the five larvae present in gallery, whereas the female (FV0994) was considered a parent (see Tables 2 and 3).
Across all Fairview samples, CERVUS identified a number of parental cross-assignments among the adult beetles present in the 15 galleries containing adults and larvae. For example, females FV0694 (gallery 3-1-58) and FV0994 (gallery 5-5-166) were assigned as the mothers of larvae FV0916 (gallery 5-2-135) and FV0864 (gallery 5-1-132), respectively. Males FV0928 (gallery 5-2-141) and FV1006 (gallery 5-5-169) were assigned as the fathers of larvae FV0851 (4-5-131), and FV0901 and FV0906 (both from 5-1-134), respectively (see Tables 2 and 3). The probability of identity from CERVUS indicated that all individuals had unique genotypes (probability of identity=0).
Using all available Fairview samples, we assessed the occurrence of full and half-sibling relatedness within galleries, and among galleries within sections, sections within trees and among trees. CERVUS showed that in a single gallery, larvae can be full-siblings, half-siblings or unrelated (see Table 4). A number of larvae present in different trees were identified as being full-siblings, although a large proportion of individuals were unrelated (89.54%). The occurrence of half-siblings was greatest among different trees, followed by larvae within the same gallery. SPAGeDi indicated that 5.69% of larvae were the product of pre-emergence mating between full-siblings (r ⩾0.625). Four adult–larva combinations were highly inbred according to the pairwise relatedness estimates obtained (>0.70). Overall, the average female reproductive success was 2.89 (s.e.±0.39) larva, whereas the male success was 4.73 (s.e.±1.15).
Discussion
Fine-scale spatial genetic structure
Previous studies investigating the genetic structure of D. ponderosae have focused on the landscape scale and typically reported weak spatial genetic structure resulting in partitioning of northern and southern sampling sites, irrespective of the spatial extent of sampling (Mock et al., 2007; James et al., 2011, 2014; Samarasekera et al., 2012). However, these results did not indicate whether D. ponderosae populations were exhibiting fine-scale spatial genetic structure. The extent and location of habitat patches for D. ponderosae, typically stands of 60–160 years in lodgepole pine (Shore and Safranyik, 1992; Aukema et al., 2006), are spatially and temporally dynamic in context because of stand aging, harvesting patterns and natural disturbances (especially wildfire). Changes in availability of suitable habitat or large changes in D. ponderosae population densities may cause the species to change its dispersal behavior or mating system (Row et al., 2010), ultimately affecting genetic structure. Although changes in genetic structure may not be detected at the landscape scale because of a homogenizing effect from long-distance dispersal (Aukema et al., 2006; Safranyik et al., 2010), they may be observed at the stand scale (Pierson et al., 2013).
Our study revealed no evidence of fine-scale spatial genetic structure at the stand level. Sampling of Fairview was conducted in 2008 at the height of the epidemic phase in an area where D. ponderosae had not been recorded before 2006. We had originally believed that 2008 attacks in this area would have come from beetles dispersing short distances as the majority of insect invasions are characterized by occasional long-distance dispersal (Liebhold and Tobin, 2008). For example, Turchin and Thoeny (1993) observed ∼50% of mark-recapture southern pine beetle (Dendroctonus frontalis Zimmerman) dispersing <600 m. Once an area is colonized, fine-scale genetic structure can develop during the time the area is occupied as a result of family groups and continued limited dispersal, and subsequently gene flow (Jacquemyn et al., 2006; Pierson et al., 2013). Thus, we believed the Fairview stand would be colonized via localized migration, providing a signal of fine-scale spatial genetic structure. However, our results suggest that homogenization, as a result of long-distance dispersal, may also influence fine-scale spatial genetic structure through the contribution and combination of different genotypes from multiple source locations, similar to the pattern observed by Janes et al. (2014). Similar conclusions have been made in locusts (Locusta migratoria), although in this instance it was suggested that the propensity to outbreak also played a major role in homogenizing genetic variation (Chapuis et al., 2009).
Individuals from Fairview appeared to be more closely related to individuals from Fox Creek, ∼700 km away. However, it is not clear whether this pattern is an artifact of temporal differences in sampling and population establishment times. Fort St James, which was sampled earlier (2005) and had been established longer than either Fox Creek or Fairview, was more unrelated to Fairview than expected. These results suggest that Fox Creek, not Fort St James or Grande Prairie, was potentially the source population for Fairview—or vice versa. Our results are similar to those found in D. frontalis. Schrey et al. (2008) compared epidemic populations of D. frontalis across national forests in Mississippi, incorporating a study area of <500 km2. They did not observe significant spatial genetic structure at this scale and proposed that the beetles in the outbreak area comprised a single interbreeding population in which genetic differentiation had not yet occurred. These results are also consistent with findings from Blanchett et al. (2012), in which rates of genetic differentiation were found to be lower in outbreak locust species compared with other Calliptamus species. We were unable to find previous studies of fine-scale spatial genetic structure in D. ponderosae.
Mating system
Mating behavior in which males or females have more than one mate at a time (that is, polygamy) is believed to evolve when the environmental potential for multiple mates is energetically defendable and individuals have the ability to utilize such potential (Emlen and Oring, 1977; Ptak and Lachmann, 2003). Polygyny is hypothesized to evolve when environment or behavior brings about female clumping, an abundance of female mates during a limited period of time, providing an opportunity to increase overall fitness (Emlen and Oring, 1977; Baena and Macias-Ordonez, 2015). Similarly, polyandry is believed to provide genetic benefits such as improving the likelihood that a female will acquire ‘good’ genes, increase genetic diversity among offspring and ensure fertilization if some males have low-quality sperm (Yasui, 1998; Bird et al., 2012). Using molecular methods we directly assessed the likelihood of polygyny vs polyandry within the Fairview D. ponderosae population.
Previous empirical evidence had proposed a mating system where males are polygynous and females are monogamous (Reid, 1958, 1962). Males were assumed polygynous because, under natural conditions, several authors indicate that a male beetle will occasionally leave a mated female and her egg gallery to mate with another (Reid, 1958, 1962; Safranyik and Wilson, 2006; Bleiker et al., 2013). Females were thought to be monogamous from laboratory observations that: (1) females can create a second egg gallery and brood in the absence of a second mating (Reid, 1958); and (2) because the male beetle arriving in the egg gallery blocks the entrance after mating, first with his body and eventually with boring dust and frass (Safranyik and Wilson, 2006). Thus, it was assumed that there was no biological need for further mating, with respect to females. However, our data clearly show, through the identification of cross-gallery and cross-tree parental assignments, that polyandry occurs under natural conditions. Biologically, this result is supported by the fact that a number of males leave the egg gallery before female oviposition (Safranyik and Wilson, 2006), and that a number of unmated males were apparently still searching for mates. Thus, ‘new’ males may take advantage of previously mated females after the original male has vacated the gallery. It is also possible that some females leave the gallery in search of more male mates to ensure fertilization and increasing fitness through multiple breedings, similar to the behavior observed in Leptinotarsa undecimlineata (Baena and Macias-Ordonez, 2015). Genetic confirmation of polyandry supports observations of polygamy in beetles from southwestern Alberta and adjacent British Columbia in the 1980s (DW Langor, unpublished data). Thus, the data presented herein suggest that D. ponderosae is best treated as polygamous, at least in epidemic phases.
When numerous adults are breeding in close proximity, these parents may interact with other parents and nonrelated offspring, expanding the potential network of social interactions. These between-family interactions may be beneficial, in the case of cooperative behaviors, or costly, in the case of competition or brood parasitism (Wong et al., 2013). Our study revealed a number of galleries containing unrelated individuals. Reasons for this may include: (1) misidentified samples, (2) larval coalescence or (3) brood parasitism. Sampling errors are unlikely as every precaution was taken to ensure errors did not occur. At high brood densities, related pupae may coalesce in a common feeding chamber before emerging (Bleiker et al., 2013). However, larval behavior may be different because of the risk of competition and subsequent density-dependent mortality. Amman and Cole (1983) state that larvae within a brood gallery will occasionally cross sibling feeding galleries, whereas Safranyik and Wilson (2006) state that larvae will rarely cross nonrelated galleries, preferring to backtrack and feed in another direction. Larvae crossing into nonrelated galleries may occur but we did not detect larval coalescence during this study.
Another explanation for the high rate of nonrelated individuals within galleries may be brood parasitism. Several insects are known to engage in brood parasitism, ranging from bees and wasps (Hymenoptera; Field, 1992) to lace bugs (Hemiptera; Zink, 2000) and beetles (Coleoptera; Müller and Eggert, 1990). Brood parasitic strategies are thought to evolve when breeding sites are in close proximity and there is an opportunity for parental care to be misdirected (Wong et al., 2013). Although brood parasitism has not been reported in Dendroctonus it is possible that, under epidemic conditions, female D. ponderosae may engage in such behavior. Brood parasitism is known to alleviate some of the energetic costs involved with parental care by making use of unrelated conspecifics (Field, 1992). It can also be initiated when breeding sites are limited as it may increase fecundity of the parasitic female in relation to the primary (Zink, 2000). Under epidemic conditions, competition among females for access to gallery space may be high, even if density-dependent competition is somewhat regulated by the cessation of aggregation pheromone production (Safranyik and Wilson, 2006). For example, endemic populations of D. ponderosae typically display a density of 7–13 individual attacks per tree (Carroll et al., 2006) with females constructing galleries up to 1.5 m in length (Reid, 1962). Under epidemic conditions the attack rate increases to ∼60 individuals per m2 (Raffa and Berryman, 1983). In such a competitive environment, it may make biological sense for some females to become ‘parasitic’ and enter another’s gallery, thereby trading valuable energetic resources normally spent on gallery construction and parental care (for example, removing frass from the galleries and remaining within the gallery after egg laying) for greater egg-laying and reproductive fitness. Brood parasitism, or sharing, cannot be confirmed by our data alone, though it is consistent with earlier anecdotal observations of brood parasitism by one of us (DW Langor, unpublished data). For the current study, the probability of multiple individuals possessing the same genotype (probability of identity) was effectively zero, making brood parasitism a plausible explanation for the presence of nonrelated larvae within galleries.
Pre-emergence mating among siblings has been reported in the ranges of 1–2% (Safranyik and Wilson, 2006) and 3–12% (Bleiker et al., 2013) for D. ponderosae. Field-caught females with full spermathecal sacs (Reid, 1958; Bleiker et al., 2013) are presumably the result of sibling crosses as only related males should be present with newly emerging females at that time. Using molecular methods in an epidemic population, our results suggest that the number of individuals resulting from matings among siblings is comparable (5%) to those previously reported. Levels of pre-emergence mating may be higher in endemic populations when density of beetles may be lower. At higher densities, there will be greater mate choice and opportunity compared with periods of low population density. Therefore, it may be more common for pre-emergence mating to occur during endemic phases. The confirmed individuals with relatedness estimates well within the range of offspring resulting from sibling matings further corroborates pre-emergence mating behavior.
The identification of highly related individuals, and the identification of half-siblings in the same gallery, adds support to a polyandrous mating system as it suggests that the pre-emergence mated females were later joined by an unrelated male in the egg gallery, resulting in two sources of sperm. We have made several anecdotal observations of two males being present in a single gallery in both field and laboratory studies (DW Langor, unpublished data). Until now, there appears to have been no published record of this behavior in D. ponderosae, although a genetic study of Dendroctonus micans Kugelann has revealed polyandry through multiple paternities in single-female broods (Fraser et al., 2014). We identified two male beetles in the same gallery (5-5-170) and genetically confirmed that both males had sired offspring present within the gallery. This finding changes our understanding of mountain pine beetle mating behavior and confirms previous unpublished observations.
Conclusion
In summary, we have confirmed that fine-scale spatial genetic structure is absent from D. ponderosae populations and that the mating system is most likely polygamous. Furthermore, we have demonstrated the effectiveness of high-throughput molecular markers in assessing population structure and their applicability in parentage studies of forest insect pests. These results have important implications in the management of current and future D. ponderosae outbreaks. A polygamous mating system, coupled with long-distance dispersal, effectively dilutes any signal of fine-scale spatial genetic structure through considerable genetic heterogeneity. The absence of fine-scale genetic structure at a localized scale implies a level of genetic uniformity (that is, each stand will exhibit a high degree of genetic variation), suggesting that any attempts to tailor forest management practices to a specific genetic component will be fruitless. This notion corroborates the idea put forth by Schrey et al. (2008) that management practices designed for one forest stand will be suitable across a broader geographic area. Our results suggest that current proactive forest management practices in which stand age cohorts are thinned by the removal of trees meeting a specific diameter at breast height, and reactive management in which ‘red’ trees are identified and removed, are suitable for managing mountain pine beetle on the landscape. It seems appropriate, given this information, that more investment should be placed on managing beetle outbreaks on a landscape scale and that the ongoing research of the population dynamics and epidemiology of D. ponderosae is key to developing effective long-term management of this pest.
Data archiving
Genotype data available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.kb418.
Change history
09 December 2015
This article has been corrected since Advance Online Publication and a corrigendum is also printed in this issue.
References
Amman GD, Cole WE . (1983). Mountain pine beetle dynamics in lodgepole pine forests. Part II: population dynamics. USDA Forest Service, Intermountain Forest and Range Experiment Station. General Technical Report INT-145.
Aukema BH, Carroll AL, Zu J, Raffa KF, Sickley TA et al. (2006). Landscape level analysis of mountain pine beetle in British Columbia, Canada: spatiotemporal development and spatial synchrony within the present outbreak. Ecography 29: 427–441.
Aukema BH, Carroll AL, Zheng Y, Zhu J, Raffa KF, Moore D et al. (2008). Movement of outbreak populations of mountain pine beetle: influences of spatiotemporal patterns and climate. Ecography 31: 348–358.
Baena ML, Macias-Ordonez R . (2015). Mobility and mating frequency in the scramble competition polygyny of a chrysomelid beetle. Behav Ecol 26: 416–424.
Bentz B, Vandygriff J, Jensen C, Coleman T, Maoney P, Smith S et al. (2014). Mountain pine beetle voltinism and life history characteristics across latitudinal and elevational gradients in the western United States. Forest Sci 60: 434–449.
Berger-Wolf TY, Sheikh SI, DasGupta B, Ashley MV, Caballero IC, Chaovalitwongse W et al. (2007). Reconstructing sibling relationships in wild populations. Bioinformatics 23: i49–i56.
Bird KL, Aldridge CL, Carpenter JE, Paszkowski CA, Boyce MA, Coltman DW . (2012). The secret sex lives of sage-grouse: multiple paternity and intraspecific nest parasitism revealed through genetic analysis. Behav Ecol 24: 29–38.
Blanchet E, Lecoq M, Sword GA, Pages C, Blondin L, Billot C et al. (2012). Population structures of three Calliptamus spp. (Orthoptera: Acrididae) across the Western Mediterranean Basin. Eur J Entomol 109: 445–455.
Bleiker KP, Heron RJ, Braithwaite EC, Smith GD . (2013). Pre-emergence mating in the mass-attacking bark beetle, Dendroctonus ponderosae (Coleoptera: Curculionidae). Can Entomol 145: 1–8.
Carroll AL, Aukema BH, Raffa KF, Linton DA, Smith GD, Lindgren BS . (2006). Mountain Pine Beetle outbreak development: the endemic-incipient epidemic transition. Canadian Forest Service, Mountain Pine Beetle Initiative Project 1.03.
Chapuis MP, Loiseau MA, Michalakis Y, Lecoq M, Franc A, Estuop A . (2009). Outbreaks, gene flow and effective population size in the migratory locust, Locusta migratoria: a regional-scale comparative survey. Mol Ecol 18: 792–800.
Cullingham CI, Cooke JEK, Dang S, Davis CS, Cooke BJ, Coltman DW . (2011). Mountain pine beetle host-range expansion threatens boreal forest. Mol Ecol 20: 2157–2171.
Emlen ST, Oring LW . (1977). Ecology, sexual selection, and the evolution of mating systems. Science 197: 215–223.
Fayt P, Machmer MM, Steeger C . (2005). Regulation of spruce bark beetles by woodpeckers — a literature review. For Ecol Manag 206: 1–14.
Field J . (1992). Intraspecifc parasitism as an alternative reproductive tactic in nest-building wasps and bees. Biol Rev 67: 79–126.
Fraser CI, Brahy O, Mardulyn P, Dohet L, Mayer F, Gregoire JC . (2014). Flying the nest: male dispersal and multiple paternity enables extrafamilial matings for the invasive bark beetle Dendroctonus micans. Heredity 113: 327–333.
Hardy OJ, Vekemans X . (2002). SPAGeDi: a versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol Ecol Notes 2: 618–620.
Holm S . (1979). A simple sequentially rejective multiple test procedure. Scand J Stat 6: 65–70.
Jacquemyn H, Brys R, Vandepitte K, Honnay O, Roldan-Ruiz I . (2006). Fine-scale genetic structure of life history stages in the food-deceptive orchid Orchis purpurea. Mol Ecol 15: 2801–2808.
James PMA, Coltman DW, Murray BW, Hamelin RC, Sperling FAH . (2011). Spatial genetic structure of a symbiotic beetle-fungal system: toward multi-taxa integrated landscape genetics. PLoS One 6: e25359.
Janes JK, Li Y, Keeling CI, Yuen MMS, Boone CK, Cooke JEK et al. (2014). How the mountain pine beetle (Dendroctonus ponderosae) breached the Canadian Rocky Mountains. Mol Biol Evol 31: 1803–1815.
Kalinowski ST, Taper ML, Marshall TC . (2007). Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol Ecol 16: 1099–1106.
Lee S, Kim J, Breuil C . (2006). Diversity of fungi associated with the mountain pine beetle, Dendroctonus ponderosae and infested lodgepole pines in British Columbia. Fungal Divers 22: 91–105.
Liebhold AM, Tobin PC . (2008). Population ecology of insect invasions and their management. Annu Rev Entomol 53: 387–408.
Maness H, Kushner PJ, Fung I . (2012). Summertime climate response to mountain pine beetle disturbance in British Columbia. Nat Geosci 6: 65–70.
Mock KE, Bentz BJ, O'Neill EM, Chong JP, Orwin J, Pfrender ME . (2007). Landscape-scale variation in a forest outbreak species, the mountain pine beetle (Dendroctonus ponderosae. Mol Ecol 16: 553–568.
Müller JK, Eggert A . (1990). Time-dependent shifts between infanticidal and parental behavior in female burying beetles: a mechanism of indirect mother-offspring recognition. Behav Ecol Sociobiol 27: 11–16.
Myrholm CL, Langor DW . (2015). Assessment of the impact of symbiont Ophiostomatales (Fungi) on mountain pine beetle (Coleoptera: Curculionidae) performance on a jack pine (Pinaceae) diet using a novel in vitro rearing method. Can Entomol e-pub ahead of print 15 June 2015 doi:10.4039/tce.2015.28.
Natural Resources Canada. (2015a). Mountain pine beetle (factsheet). Available at: http://www.nrcan.gc.ca/forests/insects-diseases/13397 (last accessed 5 March 2015).
Natural Resources Canada. (2015b). Mountain pine beetle. The threat of mountain pine beetle to Canada’s boreal forest. Available at: http://www.nrcan.gc.ca/forests/insects-diseases/13381 (last accessed 5 March 2015).
Peakall R, Smouse PE . (2006). GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Resour 6: 288–295.
Pierson JC, Allendorf FW, Drapeau P, Schwartz MK . (2013). Breed locally, disperse globally: fine-scale genetic structure despite landscape-scale panmixia in a fire-specialist. PLoS One 8: e67248.
Ptak SE, Lachmann M . (2003). On the evolution of polygyny: a theoretical examination of the polygyny threshold model. Behav Ecol 14: 201–211.
Raffa KF, Berryman AA . (1983). The role of host plant resistance in the colonization behavior and ecology of bark beetles (Coleoptera: Scolytidae). Ecol Monogr 53: 27–49.
Rankin LJ . (1988) Competitive interactions between the mountain pine beetle and the pine engraver in lodgepole pine. Master of Pest Management Professional Paper. Simon Fraser University: Burnaby, British Columbia, 33 p.
Reeve JD, Anderson FE, Kelley ST . (2012). Ancestral state reconstruction for Dendroctonus bark beetles: evolution of a tree killer. Environ Entomol 41: 433–730.
Régnière J, Bentz B . (2007). Modeling cold tolerance in the mountain pine beetle, Dendroctonus ponderodae. J Insect Physiol 53: 559–572.
Reid RW . (1958). The behaviour of the Mountain Pine Beetle, Dendroctonus monticolae Hopk., during mating, egg laying and gallery construction. Can Entomol 90: 505–509.
Reid RW . (1962). Biology of the mountain pine beetle, Dendroctonus monticolae Hopkins, in east Kootenay region of British Columbia II. Behaviour in the host, fecundity, and internal changes in the female. Can Entomol 94: 605–613.
Rice AV, Thormann MN, Langor DW . (2008). Mountain pine beetle-associated blue-stain fungi are differentially adapted to boreal temperatures. Forest Pathol 38: 113–123.
Rousset F . (2008). GenePop '007: a complete reimplementation of the GenePop software for Windows and Linux. Mol Ecol Resour 8: 103–106.
Row JR, Blouin-Demers G, Lougheed SC . (2010). Habitat distribution influences dispersal and fine-scale genetic population structure of eastern foxsnakes (Mintonius gloydi across a fragmented landscape. Mol Ecol 19: 5157–5171.
Safranyik L, Carroll AL, Regniere J, Langor DW, Reil WG, Shore TL et al. (2010). Potential for range expansion of mountain pine beetle into the boreal forest of North America. Can Entomol 142: 415–441.
Safranyik L, Wilson B . (2006) The mountain pine beetle: a synthesis of biology, management, and impacts on lodgepole pine. Natural Resources Canada: Victoria, British Columbia.
Samarasekera GDN, Bartell NV, Lindgren BS, Cooke JEK, Davis CS, James PMA et al. (2012). Spatial genetic structure of the mountain pine beetle (Dendroctonus ponderosae) outbreak in western Canada: Historical patterns and contemporary dispersal. Mol Ecol 21: 2931–2948.
Schrey NM, Schrey AW, Heist EJ, Reeve JD . (2008). Fine-scale genetic population structure of Southern Pine Beetle (Coleoptera: Curculionidae) in Mississippi forests. Environ Entomol 37: 271–276.
Sequenom. (2008). iPlex Gold Assay for SNP genotyping. BioTechniques Protocol Guide, 37 p.
Shore TL, Safranyik L . (1992). Susceptibility and risk rating systems for the mountain pine beetle in lodgepole pine stands. Victoria, British Columbia. Forestry Canada, Pacific and Yukon Region, Pacific Forestry Centre. Information Report BC-X-336.
Sturgeon KB, Mitton JB . (1986). Allozyme and morphological differentiation of mountain pine beetles Dendroctonus ponderosae Hopkins (Coleoptera: Scolytidae) associated with host tree. Evolution 40: 290–302.
Taylor SW, Carroll AL . (2004). Disturbance, forest age, and mountain pine beetle outbreak dynamics in BC: a historical perspective. Victoria, British Columbia, Canadian Forest Service, Pacific Forestry Centre. Information Report BC-X-399.
Trzcinski MK, Reid ML . (2009). Intrinsic and extrinsic determinants of mountain pine beetle population growth. Agric Forest Entomol 11: 185–196.
Turchin P, Thoeny WT . (1993). Quantifying dispersal of southern pine beetles with mark-recapture experiments and a diffusion model. Ecol Appl 3: 187–198.
Wang J, Santure AW . (2009). Parentage and sibship inference from multilocus genotype data under polygamy. Genetics 181: 1579–1594.
Wong JWY, Meunier J, Kolliker M . (2013). The evolution of parental care in insects: the roles of ecology, life history and the social environment. Ecol Entomol 38: 123–137.
Yasui Y . (1998). The "genetic benefits" of female multiple mating re-considered. Trends Ecol Evol 13: 246–250.
Zink AG . (2000). The evolution of intraspecfic brood parasitism in birds and insects. Am Nat 155: 395–405.
Acknowledgements
We acknowledge funding for this research under the Tria 1 and Tria 2 Projects (http://www.thetriaproject.ca) from Genome Canada, the Government of Alberta through Genome Alberta and the Government of British Columbia through Genome British Columbia; and from Alberta Innovates Bio Solutions. We are indebted to numerous personnel from the University of Alberta and Alberta Environment and Sustainable Resource Development for sample collection, McGill University and Genome Quebec Innovation Center for genotyping and Matthew Bryman for project management.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on Heredity website
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Janes, J., Roe, A., Rice, A. et al. Polygamy and an absence of fine-scale structure in Dendroctonus ponderosae (Hopk.) (Coleoptera: Curcilionidae) confirmed using molecular markers. Heredity 116, 68–74 (2016). https://doi.org/10.1038/hdy.2015.71
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/hdy.2015.71
This article is cited by
-
Fine-scale invasion genetics of the quarantine pest, Anoplophora glabripennis, reconstructed in single outbreaks
Scientific Reports (2019)
-
Biosurveillance of forest insects: part I—integration and application of genomic tools to the surveillance of non-native forest insects
Journal of Pest Science (2019)
