
Abstract
Over the last 5 years, disorders of nonmotile cilia have come of age and their study has contributed immeasurably to our understanding of cell biology and human genetics. This review summarizes the main features of the ciliopathies, their underlying genetics, and the functions of the proteins involved. We describe some of the key findings in the field, including new animal models, the role of ciliopathy proteins in signaling pathways and development, and the unusual genetics of these diseases. We also discuss the therapeutic potential for these diseases and finally, discuss important future work that will extend our understanding of this fascinating organelle and its associated pathologies.
Similar content being viewed by others

THE MEDICAL AND SCIENTIFIC IMPORTANCE OF THE CILIOPATHIES
The ciliopathies form a class of genetic disease whose etiology lies with primary ciliary dysfunction. Although many of these syndromes have been recognized as isolated entities, it has only been in the last few years that this seemingly disparate collection of rare and clinically perplexing disorders has been classified.
The term “nonmotile ciliopathy” refers to diseases that tend only to affect the function of nonmotile, primary cilia, and not the motile cilia, familiarly lining epithelia and powering sperm movement. Many disease gene products localize to the basal bodies and centrosomes too, but here we only refer to those conditions where the phenotype can be ascribed to aberrant cilia function or signaling. The list of bona fide ciliopathies continues to grow and at present includes Bardet-Biedl syndrome (BBS), nephronophthisis (NPHP), Senior-Löken syndrome (SNLS), Alström syndrome (ALMS), Meckel syndrome (MKS), Joubert syndrome (JBTS), Oral-facial-digital Type I (OFD 1), Jeune asphyxiating thoracic dystrophy (JATD), Ellis van Creveld (EVC), and Leber congenital amaurosis (LCA). Polycystic kidney diseases (PKD) both the recessive and dominant forms should also technically be regarded ciliopathies. In view of the existing and extensive literature on these disorders, they will not be discussed here and the reader is referred to relevant reviews.1,2
THE IMPORTANCE OF UNDERSTANDING CILIARY DISEASE
Despite their relative rarity (incidence of individual ciliopathies ranges from around 1 in 1000 [for autosomal dominant polycystic kidney disease (ADPKD)] to 1 in 150,000), the study of ciliopathies has given rise to an important and dynamic new field of biology, which in turn is revealing previously unrecognized cellular phenomena. Understanding these diseases has been helped enormously by basic research into ciliated organisms such as the unicellular alga Chlamydomonas reinhardtii and the flat-worm, Caenorhabditis elegans. The ciliopathies have, in return, shed light on the functional importance of cilia and their proteins in signaling and development. As such, the study of ciliopathies is a perfect example of how basic science can be used to understand human disease, and vice versa.
The ciliopathies cause multisystem pathology, resulting in very low quality of life and early death for many patients. The availability of molecular diagnosis to these patients and their families is therefore crucial for determining prognosis, proactive medical, and educational management and in the case of life-shortening disorders, will assist in end-of-life planning. The availability of mutation testing will also facilitate accurate genetic counseling and antenatal screening for those at risk. Currently, there are no cures and few treatments, and so understanding disease pathogenesis is important for developing potential therapies. Although rare in their own right, several features of ciliopathies (e.g., neural tube defects [NTD], orofacial clefting) are common in the general population. Obesity is present in around one third of adults, kidney cysts in about 1 in 500, retinal degeneration around 1 in 3000, and polydactyly 1 in 500. One of the most prominent features of BBS is obesity, present in up to one third of adult Britons. In BBS, this is combined with Type 2 diabetes, hypertension, and cardiovascular disease, and so provides a good model for studying the poorly understood metabolic syndrome, which affects around 25% of the American population. We suggest that careful scrutiny of the ciliopathies may provide unique insights into more common medical problems.
THE FEATURES AND COMMON DENOMINATORS OF CILIOPATHIES
The motile ciliopathies (such as primary ciliary dyskinesia) are clinically distinct from the nonmotile ciliopathies. Despite both classes of diseases affecting a structurally similar organelle, because of the differing biology of motile and primary cilia, the clinical involvement differs. Because of their common involvement in ciliary processes, the ciliopathies display common clinical features. In fact, these features are so characteristic that they can be used as a means to predict and identify new ciliopathy syndromes.3 This can then be used to discover new ciliary proteins and further our understanding of ciliopathy. The common features of ciliopathy are presented in Table 1.
It is clear that cystic kidneys and liver disease are common to all ciliopathies. Second to this in prevalence is situs inversus, the reversal of organ laterality. Polydactyly, agenesis of the corpus callosum, and mental retardation are a triad of features that tend to manifest together. This possibly reflects common underlying signaling defects for which the cilium acts as a nexus. Retinal degeneration is common to several ciliopathies and usually culminates in blindness. Encephalocoele is a NTD in which portions of the anterior neural tube fail to close, resulting in brain protruding through the skull, and lethality.
NPHP has less multisystem involvement than the other diseases and is considered phenotypically milder than BBS or MKS. It primarily involves the kidney and is a common cause of kidney failure among children.4 When NPHP presents with retinal degeneration, it is called SNLS. BBS involves more organ systems and has become the most studied ciliopathy, partly because it is informative about the role of cilia in so many different systems, and manifests a strong phenotype without significant early lethality. By contrast, MKS and JBTS are both severe disorders, the former culminating in perinatal death. A characteristic presentation of JBTS is ataxia with cerebellar vermis hypoaplasia/aplasia. The so-called “molar tooth” sign is usually seen on magnetic resonance imaging of the posterior brain. MKS is associated with NTDs and usually presents with an encephalocoele in addition to polydactyly and renal cystic disease.
Other ciliopathies affect myriad organs, whereas preserving the hallmarks such as kidney disease, situs inversus, and polydactyly. OFD1 and JATD both cause skeletal defects such as shortened long bones and ribs. LCA only affects the retina and so blindness is the main feature.
SUBCLASSES OF CILIOPATHY
As more diseases emerge as nonmotile ciliopathies, it is becoming possible to categorize the diseases based on the clinical features of the disease, presumably contingent on the biological role of the protein involved. Excluding kidney diseases, this classification defines two groups: ciliopathies with skeletal involvement (JATD, OFD1, EVC) and those without (BBS, NPHP, MKS, JBTS, ALMS, LCA). Comprehensive description of the tissue and subcellular expression domains of these different proteins will likely enable explanation and prediction of disease features.
Bardet-Biedl syndrome
BBS has become a model ciliopathy because it became the first disease whose etiology lay in primary ciliary disorder.5 Patients with BBS have all the common features of ciliopathies: polydactyly, cystic kidneys, retinitis pigmentosa (RP), and situs inversus (Fig. 2,A and B). Nearly, all patients with BBS are obese (98% according to Beales et al., 1999), the precise reason is unclear but may be linked to a defect in the satiety center of the hypothalamus, whose neurons are ciliated.6,7

A variety of clinical features associated with nonmotile ciliopathies. A, Fundoscopic image demonstrating retinitis pigmentosa and cataract in a 32-year-old man with Bardet-Biedl syndrome. B, Postaxial polydactyly of the left foot in an 8-year child with Bardet-Biedl syndrome. C, Transverse T1-weighted MR image obtained at the level of the midbrain reveals thickened superior cerebellar peduncles (arrows) with a deep midline cleft at the pontomesencephalic junction. (Reproduced with permission from: McGraw P. Radiology 2003;229:671–672). D, Radiograph showing typical narrowed thorax with short slender ribs in a child with Jeune asphyxiating thoracic dystrophy and mutations in IFT80. E, Glomerular hyalinization and fibrosis in a kidney section from a patient with Alstrom syndrome (Reproduced from: Beales et al. In The genetics of renal disease, Oxford University Press, 2003). F, Bilateral renal cystic disease in a fetus with Meckel syndrome (Jean-Christophe Fournet, Humpath.com).
Patients with BBS are usually born with postaxial polydactyly of some or all limbs and hypogenitalism. In the first few years of life, they begin to put on weight and become obese. Usually by the age of 8, night blindness manifests, proceeding to complete blindness by about 15 years. Kidney cysts often appear in childhood and some patients require dialysis or transplantation. End-stage renal failure (ESRF) is the most common cause of premature death from BBS, whereas around 30% of patients will develop chronic kidney disease.8 Complications arise from being overweight, including hypertension and Type 2 diabetes. Many patients also have congenital cardiovascular disease. To date mutations in 15 genes are associated with BBS; BBS1–12, MKS1, MKS3, and CEP290. The function of some of these proteins is discussed later.
Alström syndrome
The ALMS is a rare recessive disorder that shows strong resemblance to BBS. It is characterized by cone-rod dystrophy, neurosensory hearing loss, early-onset obesity and insulin resistance leading to Type 2 diabetes. A number of other features may also present such as dilated cardiomyopathy, hepatic and renal dysfunction (Fig. 2E), short stature, and male hypogonadism.9 Mutations in ALMS1, account for all cases of this syndrome.10,11 ALMS1 is ubiquitously expressed throughout all organ tissues10; it was first identified in a proteomic study of the centrosome12 where it is localized to the centrosome and basal body,13 which suggested involvement in ciliary function and provides a possible explanation for the phenotypic overlap with BBS.11 Studies in foz (fat aussie) mouse mutants that carry a mutation in Alms1 and an Alms1 knock-out mouse suggest that ALMS1 has a role in intracellular trafficking.14,15 These mice develop features similar to patients with ALMS such as obesity, hypogonadism, hyperinsulinemia, retinal degeneration, late-onset hearing loss and males, as in Bbs null mice, are infertile due to aflagellate spermatozoa.
Nephronophthisis
The NPHP is an autosomal recessive cystic renal condition characterized by corticomedullary clustering of cysts and tubulointerstitial fibrosis. In contrast to PKD where enlarged kidneys are a common diagnostic feature, the overall size of the kidney in NPHP is normal or diminished. Although, strictly speaking, NPHP describes a renal histopathology, ∼10% of cases also present with extrarenal manifestations that can be associated with other syndromes such as RP (SLSN), cerebellar vermis hypoplasia (JBTS), ocular motor apraxia (Cogan type), cognitive impairment, hepatic fibrosis, phalangeal cone-shaped epiphyses (Mainzer-Saldino), and situs inversus. NPHP has also been described in cases of BBS, EVC, JATD, ALMS, and MKS.4 NPHP presents as three forms characterized by the time of onset of ESRD: infantile, juvenile, and adolescent. Collectively, they constitute the most frequent genetic cause of ESRF in the young. Eight causative genes have now been identified (NPHP1–8) and the analysis of their protein products has provided a strong link between ciliary function and the pathogenesis of this disease.
Meckel syndrome
The lethal autosomal recessive MKS is characterized by renal cystic dysplasia (Fig. 2F) with fibrotic changes in the liver, occipital encephalocoele or some other central nervous system malformation. Additionally, polydactyly is frequently reported and some patients have cleft palate, cardiac abnormalities, and incomplete development of genitalia and gonads.16–19 Patients with MKS invariably die from respiratory and/or renal failure. Thus far, MKS has been linked to six loci of which five genes have so far been identified; MKS1, MKS3 –6– some of these are discussed later. All identified gene products are associated with ciliary functions.
Joubert syndrome
The JBTS is an autosomal recessive condition characterized by hypotonia, ataxia, severe psychomotor delay, oculomotor apraxia, and episodes of rapid breathing. Diagnosis may be supported by the neuroradiological hallmark referred to as the “molar tooth sign” (MTS, Fig. 2C), owing to horizontally oriented and thickened superior cerebellar peduncles and a deepened interpeduncular fossa combined with cerebellar vermis hypoplasia.20 The MTS has greatly enhanced the diagnosis of JBTS and with this has identified a group of JBTS-related disorders with involvement of other organs. For instance, approximately a quarter of patients develop juvenile NPHP and retinal dystrophy, termed cerebello-oculo-renal syndrome or JBS Type B.21 Several additional clinical features have been reported including occipital encephalocele, polymicrogyria, cystic kidneys, polydactyly, hepatic fibrosis, and ocular coloboma thus overlapping with the lethal, recessive disorder MKS and SLSN. Six of the eight JBTS loci have now been described; again a clear genetic overlap with other ciliopathies is apparent. The underlying gene mutations for JBTS1 and JBTS2 loci are not yet identified; however, JBTS3 has been shown to be caused by mutations in the AHI1 gene22,23 that accounts for between 7 and 11% of JBTS cases most of which are accompanied by retinopathy.24–26 The AHI1 gene product is Jouberin.
JBTS8/ARL13B mutations have recently been identified in families with a classical form of JBS.27 ARL13B belongs to the Ras GTPase family and in other species is required for ciliogenesis, body axis formation, and renal function. Two animal models have been reported for this gene: the lethal hnn (hennin) mouse that has a coupled defect in cilia structure and Sonic hedgehog (Shh) signaling and the zebrafish scorpion mutant that displays renal cysts and a curved tail, both of which are phenotypes common in morphants with cilia dysregulation.27–30
Jeune syndrome
The JATD is an autosomal recessive chondrodysplasia. Affected children often die in the perinatal period owing to respiratory insufficiency that is a consequence of narrow and slender ribs and abnormal cage formation (Fig. 2D). Radiographical analysis can also indicate a shortening of the long bones and changes in the pelvic bones and the phalanges. There can be multiorgan involvement such as biliary dysgenesis with portal fibrosis and bile duct proliferation, renal cystogenesis and failure, polydactyly, and retinal degeneration.
These phenotypic clues led to the identification of mutations in IFT80 in a subgroup of patients presenting with milder disease without renal, liver, pancreatic, or retinal features.3 There is, however, genetic heterogeneity with another, as yet unidentified locus on 15q. IFT80 is a member of the intraflagellar transport (IFT) complex B proteins that are important for cilia structure and function. Ift80 knock-down in the multiciliate protozoan, Tetrahymena, resulted in fewer cilia and nuclear duplication.3 In zebrafish, silencing ift80 results in convergent extension defects, cystic pronephros, and cardiac edema whereas knock-out mice all display early embryonic lethality (unpublished observations).
Ellis van Creveld syndrome
Also known as chondro-ectodermal dysplasia, EVC is characterized by short ribs, polydactyly, growth retardation, ectodermal (dysplastic finger nails and teeth), and cardiac defects.31 EVC is rare (approximately 150 cases described) and is inherited as an autosomal recessive trait with variable expression. Causative mutations in EVC1 and EVC2 genes have been identified, located in a unique head-to-head configuration on 4p16. Renal abnormalities have been described in a few cases with agenesis, dysplasia, megaureter, and nephrocalcinosis. Lethal NPHP has been reported once.
Oral-facial-digital syndrome
The OFD Type I syndrome is an X-linked dominant disease characterized by malformations of oral cavity, face, and digits and by cystic kidneys. Facial features include hypertelorism, broad nasal bridge, buccal frenulae, cleft palate, lobulated tongue, lingual hamartomas; in the hands and feet, brachydactyly and polydactyly may be present. PKD is common and central nervous system malformations include corpus callosum agenesis, cerebellar abnormalities, and hydrocephalus, with accompanying mental retardation. It is presumed lethal in males. Mutations in the novel gene OFD1 are causative and the OFD1 protein localizes both to the primary cilium and to the nucleus.32,33 Ferrante et al.34 knocked out Ofd1 in mice and recapitulated the human phenotype albeit with increased severity, possibly owing to differences of X inactivation patterns between species. They also showed a failure of left-right axis specification in mutant male embryos, a lack of cilia in the embryonic node, mispatterning of the neural tube and altered expression of Hox genes in the limb buds, all of which are indicative of cilia defects and demonstrate that Ofd1 plays a role in ciliogenesis.
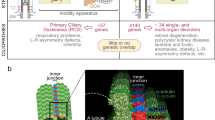
CILIARY BIOLOGY: PRIMARY VERSUS MOTILE CILIA STRUCTURE
Motile cilia are long thin protrusions, extending up to 20 μm from the cell surface. They tend to be concentrated in large numbers on the apical surface of cells and beat in coordinated waves to clear mucus from the respiratory epithelium, drive sperm along the fallopian tube, and move cerebrospinal fluid in the brain ventricles and spinal cord. In cross section, these cilia are constructed from a “9 + 2” arrangement of microtubules, in which nine microtubule doublets surround a central inner pair (Fig. 1). These microtubules are stabilized by acetylation and therefore not dynamically unstable as they are in the cytoplasm.

Schematic diagram of the primary cilium illustrating the concept of intraflagellar transport (IFT) and the component parts therein. The protein cargo is manufactured in the Golgi apparatus and carried by vesicles to the cell membrane, where receptor proteins may also be picked up and translocated to the IFT particles docked at the transition zone. Under the influence of the kinesin motors, the IFT/cargo assembly is transported along the microtubular axoneme (anterograde transport). At the tip, cargo may be unloaded, further proteins/molecules and kinesin itself is loaded and carried back down the axoneme (retrograde transport) facilitated by the dynein molecular motor. In this way proteins are destined for degradation or recycling.
Each doublet consists of an “A” strand, which is constructed from 13 tubulin protofilaments, and a “B” strand made from 10 protofilaments. In motile cilia, the outer and inner doublets are connected by radial spokes, which are used to bend the outer doublets relative to the inner, producing a sheer force necessary to bend the cilium.
Primary cilia lack the central microtubule doublet and so are described as having a “9 + 0” arrangement. The 9 + 0 cilia are immotile and do not beat, with the unusual exception of cilia covering the node (or organizer) of the vertebrate embryo. The function of primary cilia, once regarded as vestigial,35 is complex and variable. In some cells, they function as antennae to respond to extracellular signaling molecules, in others they bend in response to fluid flow, and they are also crucial for certain important developmental signaling cascades.
The ciliary axoneme is only 0.25 μm in diameter, but its length can reach up to 20 μm depending on cell type. The basal body is a cylindrical structure positioned perpendicular to the cell membrane, anchoring the cilium in the cytoplasm. It also acts as a nucleation point from which the cilium extends out into the cell. The basal body consists of nine triplets of microtubules arranged in a ring, from which extend three microtubules: A, B, and C. All three fibers project from the basal body into the proximal region of the cilium, known as the transition zone, from which a nascent cilium grows.
Intraflagellar transport
The ciliary matrix, an extension of the cytoplasm, is devoid of ribosomes, meaning that any proteins needed for ciliary biogenesis or function must be transported into the cilium from the cytoplasm. A system of ferrying proteins within the cilium either toward the tip (anterograde) and from the tip (retrograde) is employed, termed IFT. Anterograde transport involves the loading of cargo proteins bound for the cilium onto an IFT particle, which in turn is attached to a kinesin motor protein complex. The motor complex consists of the motors KIF3A and KIF3B, which slides along the axoneme.36 Retrograde transport is facilitated by the dynein-dynactin motor complex.
One of the best studied mouse models of defective IFT is the Oak Ridge Polycystic Kidney (ORPK) mouse, described 14 years ago as a model for human recessive PKD (see Lehman et al.37 for review). The ORPK mouse arose through integration of a transgene into an intron of the Ift88 gene resulting in a hypomorphic allele (Ift88Tg737Rpw). This has the effect of disrupting the expression and function of the cognate protein, polaris. The ORPK phenotype includes scruffy fur, preaxial polydactyly, growth retardation with later obesity. It was the first mammalian model to establish a connection between cystic kidney disease and ciliary dysfunction. More recently, additional features include hepatic and pancreatic ductal cysts, retinal degeneration, skeletal defects, cerebellar hypoplasia, and hydrocephalus. Many of these are observed in a wide range of human ciliopathies.
The IFT particle consists of at least 17 proteins and is divided into subcomplex A, with around six proteins (IFT43, 122A, 122B, 139, 140, 144), and B with 11 proteins (IFT20, 27, 46, 52, 57, 72, 74, 80, 81, 88, 172). These two complexes are coordinated by a bridge of accessory proteins. The precise nature of the IFT particle is not completely characterized, and its 3D structure is yet to be solved. The ciliary proteome comprises some 1000 proteins,38 many of which are directly involved with the process of IFT, and so there are likely to be multiple proteins interacting with the IFT particle subunits.
The rates of antero- and retrograde transport differ, such that retrograde is slightly faster presumably to prevent the accumulation of any particles within the cilium. When the cilium is fully extended, many IFT particles enter the cilium without cargo and keep passaging up and down the axoneme. Loading of cargo is regulated at the basal body and the complex of proteins surrounding it; the pericentriolar material (PCM). Many of these cargoes derive from vesicles budded directly off the Golgi apparatus, so it is not surprising that the cilium usually forms apically to the Golgi.39
Formation and resorption of the primary cilium is dynamic and is intrinsically linked with the cell cycle. The cilium is withdrawn before mitosis as the basal body is recruited as a microtubule-organizing center to form the mitotic spindle. Cilia tend to protrude from quiescent cells that are not cycling.
Signaling at the cilium
Several critical developmental signaling pathways require the cilium to function. The current paradigm explains the primary cilium as an “antenna” for sensing extracellular signaling molecules and organizing their cytoplasmic effectors. The following pathways have been shown, to varying degrees, to rely on the cilium: Shh, canonical Wnt, noncanonical Wnt, PDGF, and mammalian target of rapamycin (mTOR).
Evidence has amassed of the importance of the cilium in Shh signal transduction. In an ethyl-N-nitrosourea-based mutagenesis screen for embryonic patterning mutants, Huangfu et al.40 identified Ift72 and Ift88. These embryos showed reduced amounts of Ptc1 expression, a direct correlate of inhibited Shh signaling. This was confirmed by a reduction of ventral cell types in the anterior neural tube and concomitant dorsal expansion of cell types normally repressed by Shh. This indicated a loss of responsiveness to Shh stimulation. Paradoxically, the limbs were polydactylous, suggestive of an expanded Shh expression domain inducing ectopic digits.
Further work attributed this defect in Shh signaling to aberrant processing of the Gli3 transcription factor from its usually full-length activator isoform to its repressive form.41 The accumulation of the active isoform of Gli3 in IFT mutants may be what caused the excessive digits to grow.
Corbit et al.42 showed that Smoothened, the membrane receptor of the Shh ligand, localized to the cilium in response to stimulation by Shh, and that its ciliary localization was necessary for downstream signaling activity. Rohatgi et al.43 localized Patched-1, which is inhibited by Smoothened in the absence of Shh, to the cilium. They showed this localization to be antagonistic to Smo ciliary localization in the absence of Shh. In further studies, IFT was shown to be necessary for Smoothened's ciliary localization.44 Su(fu), Gli2, and Gli3 have all been identified in the cilium, and this localization is abrogated in IFT mutants. Given the phenotypes of these IFT mutants, it suggests that ciliary localization of several Shh components is essential for pathway activity. Several other ciliary components have since been shown to be essential for Shh signaling, including RPGRIP1L, a causative gene of MKS and JBTS.45
Postaxial polydactyly is a common feature of the ciliopathies, present in BBS, JATD, MKS, and JBTS. The presence of several Shh-associated phenotypes in patients with ciliopathy, including: polydactyly, external genitalia anomalies, and craniofacial defects suggest a potential Shh signaling defect in these ciliopathy mutants. When BBS proteins are knocked down in cell culture, the cells become unresponsive to administration of exogenous Shh ligand. Failure of the cells to stop cleaving active Gli3 in response to Shh results in a lack of activation of downstream target genes.46
Wnt signaling is another important pathway disrupted when components of the cilium are knocked out, and which has important ramifications for the physiology of patients with ciliopathy. The canonical Wnt pathway is involved in many contexts and employs secreted Wnt ligands, which regulate cell fate and patterning during development. Noncanonical or planar cell polarity (PCP) signaling is involved in areas of organogenesis where cells are polarized throughout an epithelial plane, such as alignment of the stereociliary bundles of the cochlea, closure of the neural tube, and orientation of the mitotic spindles of renal tubular cells.47
Simons et al.48 showed that Inversin (NPHP2) localized to the cilium and acted as a molecular switch between the canonical and noncanonical Wnt pathways. Inversin inhibits canonical Wnt signaling by facilitating the degradation of cytoplasmic Disheveled (Dvl), concomitantly inducing PCP activity. They showed that fluid flow in renal cells increased the level of Inversin in cultured kidney cells. From this they speculated that urine flow in the developing renal tubule terminated canonical Wnt signaling in favor of PCP, which then patterns the tubule to give its filtrative capacity.
In ciliopathy mutants, which have cystic and dysplastic kidneys, it is tempting to suggest that unopposed canonical Wnt signaling may suppress terminal differentiation of tubular cells, leading to cyst formation. A possible molecular mechanism for this is that Inversin fails to localize to the cilium in these mutants.
Ross et al.49 showed that BBS proteins were involved in PCP signaling. Park et al.50 confirmed these observations in two separate studies. One showed that mutations in two PCP genes, inturned and fuzzy, abrogated PCP signaling and also caused Shh-deficient phenotypes, attributed to a ciliogenesis defect. The other study showed that Disheveled, a key cytoplasmic effector of PCP signaling, was required for the polarized positioning of the basal body in epithelial cells.51 These data combined might explain why mispatterned kidney tissue is so common in patients with ciliopathy.
Two articles have directly linked canonical Wnt signaling to the primary cilium. Gerdes et al.52 showed that suppression of Bbs1, Bbs4, Bbs6, and Kif3a resulted in stabilization of β-catenin and upregulation of TCF-mediated transcriptional activity indicative of over-active canonical Wnt signaling.
Corbit et al.53 showed that the canonical pathway components β-catenin and APC localized to the cilium. They showed that Ofd1 null cells were stimulated by exogenous Wnt ligand five times more than wild-type cells. They attributed this to the absence of cilia in these cells and proposed that the cilium acts to inhibit canonical Wnt signaling, in contrast to its role in Shh transduction. This suggests that knocking out ciliary proteins relieves this brake, causing excess canonical Wnt pathway activity.
If cilia were essential for all canonical Wnt signaling, one would expect to see hyperproliferation in patients with ciliopathy of cells in all tissues maintained by Wnt signaling, such as in the gut. This is not the case and only cystic kidneys are observed. Either the importance of the cilium in Wnt signaling is context dependent, or there are cilium dependent and cilium-independent functions of Wnt signaling. A further explanation for the discrepancy could be that the induction of canonical Wnt signaling in ciliary mutants is secondary to downregulation of PCP signaling.
The mTOR is a serine/threonine protein kinase that regulates cell growth, cell proliferation, cell motility, cell survival, protein synthesis, and transcription. The mTOR seems to integrate the input from multiple upstream pathways, including insulin, IGF-1, IGF-2, and mitogens. mTOR also functions as a sensor of cellular nutrient and energy levels and redox status (reviewed by Edgar et al.54). The dysregulation of the mTOR pathway is implicated in various types of cancer. Recently, two lines of evidence point to the involvement of ciliary signaling to the mTOR pathway. First, the ciliary protein, polycystin 1 spans the membrane and its intracytoplasmic tail interacts with the tuberose sclerosis protein, tuberin, and the mTOR pathway is inappropriately activated in cyst-lining epithelial cells in patients with ADPKD and mouse models.55 Uncontrolled epithelial cell proliferation is considered an important pathogenic mechanism in tubular cyst formation. The fact that the mTOR inhibitor, rapamycin resulted in a significant reduction in polycystic kidney size indicates that PC1 has an important function in the regulation of the mTOR pathway and possibly proliferation. Second, a study of zebrafish embryos injected with a morpholino against tsc1a, led to kidney cyst formation and left-right asymmetry defects.56 Furthermore, the same study indicated that Tsc1a regulates the length of the cilium itself.
Establishment of left-right asymmetry
Situs inversus is one of the “hallmarks” of ciliopathies. It is the complete reversal of organ laterality, such that the position of the heart, liver, stomach, and other internal organs is totally inverted. Often the inversion of laterality is partial and the umbrella term for laterality defects is “heterotaxia.” This occurs at a rate of 1 in 10,000 in the general population but is frequently observed (precise figures unavailable) in patients with ciliopathy.5,57–60
Situs inversus in a patient with BBS8 mutations was responsible for identifying ciliary origins of the disease, because of the canonical role for the cilium in the breaking of embryonic symmetry early in development. In the mouse embryo, the node appears during gastrulation at the anterior end of the primitive streak. The center of the node is covered in motile cilia which beat in a clockwise direction and establish the “nodal flow,” a current across the node.61,62 This flow causes the passage of vesicles, termed “nodal vesicular parcels.”63 These contain the morphogens Shh, retinoic acid, and fibroblast growth factor. These establish a left-sided accumulation of calcium ions, which activates the nodal pathway on the left side of the embryo, imparting left-sided character.62
There are two populations of cilia in the node, one responsible for establishing the flow and the other for sensing it.64 It is unknown which population is affected in the ciliopathies. As both populations are 9 + 0 cilia, it is possible that both are affected.
In the invs mouse, so called because of its inverted laterality, Nphp2 is mutated.60 This protein functions at the node, where it is important for nodal flow.65 Other ciliopathy proteins have also been localized to the node, including Ift80 (unpublished data).
The study of laterality is being facilitated by the zebrafish, in which asymmetry is established by a ciliated organ similar to the mammalian node, called Kupffer's vesicle. This forms after gastrulation at the posterior end of the embryo and contains beating cilia, which drive fluid in a clockwise direction to establish left-right identity in the paraxial mesoderm.
Oishi et al.66 identified a gene, duboraya (dub), essential for ciliogenesis and fluid flow in Kupffer's vesicle, with resultant randomization of laterality in dub deficient embryos. They showed that its function was dependent on Frizzled-2 mediated phosphorylation, linking dub to the noncanonical Wnt pathway. Several other ciliary mutants also show laterality defects observed by aberrant sidedness of expression of left sided markers such as southpaw (spaw) or lefty, or morphological phenotypes such as mis-sided heart or gut looping. These studies show that establishment of asymmetry in fish and mammals is a homologous process reliant on the same molecules and cellular processes. They also clearly link ciliary and ciliopathy proteins to asymmetry defects.
Sensory deficits
RP involves progressive loss of vision, first manifesting as night blindness, then tunnel vision, before progressing to complete loss of sight. It is a primary feature of several ciliopathies, including BBS, ALMS, and SNLS and associated with cases of JBTS and JATD. The average age of complete blindness in patients with BBS is 15.67 The connecting cilium in the rod cells carries rhodopsin molecules into the outer segment of the rod cell. Defects in ciliary transport cause these cells to die by apoptosis resulting in progressive loss of vision.
Abd-El-Barr et al.68 showed that rhodopsin is not transported from the cell body to the outer segment in Bbs4 null mice, and that these cells subsequently die. In addition, they noticed defects in synaptic transmission from the photoreceptors to the visual neurons.
Bainbridge et al.69 provided the first evidence of effective gene therapy for retinal degeneration caused by a ciliopathy—LCA. The underlying mutation was in RPE65, which encodes a ciliary protein expressed in the retina. They were able to reconstitute the sight of three patients by injecting an adenoviral vector underneath the retina of the patients. This raises the possibility of treating the visual deficit in patients with ciliopathy using gene therapy.
Deafness is also present in several of the ciliopathies, most notably ALMS. Most patients develop sensorineural hearing loss and about 10% become profoundly deaf.70 Ross et al.49 found that half the Bbs6 null mice tested failed to respond to auditory stimuli, compared with none of the controls. They found that patients with BBS could not detect high-frequency sounds, and adolescent patients had a hearing age of elderly individuals.
Etiology of kidney pathology
In the developing kidney, cells divide rapidly to produce the branched tubules required for filtration. As the embryo develops, urine begins to flow through the tubules as the kidneys begin to function as organs of excretion and osmoregulation. The onset of flow is thought to reduce cell proliferation and begin to induce terminal differentiation of tubular cells. This change of fate coincides with a shifting emphasis from canonical to noncanonical Wnt signaling.2 Primary cilia projecting from tubular epithelial cells into the lumen bend as fluid flows over them.1 This bending opens calcium channels in the membrane and causes a flux of intracellular calcium as cytoplasmic stores of calcium are released by calcium-induced calcium release.
Li et al.71 placed inner medullary collecting duct 3 cells in a flow chamber and passed a fluid flow over the cells. They found that cells expressing siRNA against ALMS1, the causative gene for ALMS (blindness, deafness, obesity, diabetes, renal dysplasia) (Fig. 2E), failed to produce a calcium spike, whereas calcium was released in control cells. The kidneys of knockout mice were cystic and had increased levels of proliferation and apoptosis, which resulted in impaired function, evidenced by proteinurea.
Kidney disease is one of the most prevalent features of ciliopathies and is the most common cause of morbidity and mortality. PKD affects between 1 in 500 and 1 in 1000 people and results in the growth of large cystic lesions, mainly in the loop of Henle, which impair the ability of the kidney to filter the blood. This can result in chronic renal failure. Currently, the only treatments are dialysis, which is expensive and not without concomitant medical problems; and transplantation, currently limited by the availability of organs and the risk of rejection. Several drugs inhibiting cystogenesis are in preclinical and clinical development.
Obesity
A potentially controversial aspect of the ciliopathic phenotype is obesity. It is one of the primary features of two ciliopathies whose diagnosis is often confused: BBS and ALMS. The frequency of being overweight in BBS varies by study from 7267 to 91%.72 The differences likely reflect different criteria for classifying obesity. Beales et al.70 found that 52% of patients had a body mass index (BMI) of over 30 (classified as obese) and 16% had a BMI >40 (morbidly obese). Early-onset accumulation of adipose tissue tends to be distributed around the trunk and hips. Childhood obesity is present in 95% of patients with ALMS.9 Therefore, obesity is a common thread among certain ciliopathies, but its precise cause is not clear.
One of the causes of obesity in BBS is thought to arise from hyperphagia (overeating). This may stem from a defect in the satiety center of the hypothalamus, in which ciliated neurons regulate the feeling of satiety and hunger, and control feeding behavior accordingly.73 Anecdotal reports claim that patients with BBS rarely feel full, even after a large meal, and so cannot avoid overeating. Strict caloric restriction is required to prevent morbid obesity ensuing.
Grace et al.74 found no difference in energy metabolism between patients with BBS and controls matched for BMI, although patients tended to be much less physically active, likely contributing to their overweight. Studies of mouse models of BBS have indicated that they consistently eat more than their wild-type littermates and develop obesity progressively from an early onset. Rahmouni et al.75 found BBS2, 4, and 6 null mice to have low locomotor activity and increased levels of circulating leptin, a hormone that regulates satiety. Adding exogenous leptin to wild-type mice reduced bodyweight by around 10% in a 4-day period, but it had no effect on BBS mutant mice, regardless of underlying mutation. This suggested that BBS mice were leptin-resistant and the hypothalamic neurons were not responding to the signal, resulting in uncontrolled appetite and a persistent feeling of hunger. They also showed that this leptin resistance contributed to hypertension in the mutant mice, another feature of BBS.
A naturally occurring mouse mutant of Alms1 exists known as Fat aussie (Foz).15 This mouse also had hyperphagia and obesity, as well as insulin insensitivity resulting in Type 2 diabetes. In addition, the sperm of males were aflagellate, causing infertility, as is the case for BBS null mice. The subcellular localization of Alms1 reveals that it has a ciliary and basal body distribution, although fibroblasts from affected individuals show no defects in cilia formation or the cytoskeleton.13
Berbari et al.7 found that G-protein coupled receptors failed to localize to primary cilia in BBS mutant mice. This included melanin concentrating hormone receptor 1, a protein involved in the regulation of feeding behavior. They suggested that this could result in hyperphagia-induced obesity.
Romano et al.76 found that Alms1 was expressed at highest levels in preadipocytes, but treating adipocytes with adipogenic factors had no effect on the gene expression of Alms1. This suggested that Alms1 might have a role in the transition from preadipocytes to adipocytes. Forti et al.77 showed that BBS1–9 and BBS11 were expressed in adipocytes, and that transcript levels increased two- to fourfold during differentiation of adipocytes.
Davenport et al.73 provided the first potential cell biological insight into the mechanism of ciliopathic obesity. They knocked out Kif3a, thereby completely ablating cilia, specifically in hypothalamic neurons expressing POMC (proopiomelanocortin), a type of neuron negatively regulating appetite. They found that these mice had one specific phenotype: uncontrolled eating leading to obesity. Although BBS knockout does not have as dramatic an effect on ciliogenesis as Kif3a, it is clear that cilia are important in this class of neuron for appetite regulation, and suggests a potential role for BBS and ALMS proteins in these neurons. Currently, no conditional mouse models exist for BBS in which this can be tested.
There seem to be potential roles for BBS proteins in appetite sensation and fat storage. The data supporting a role for the proteins in the hypothalamus are more extensive and convincing, and fit with the observation of overeating.
Skeletal abnormalities
Earlier, we classified the ciliopathies into those with or without skeletal involvement; the former group consisting of JATD, EVC, and OFD1. JATD and EVC show similar skeletal and extraskeletal features, as does short-rib polydactyly syndrome, although the gene for this has not yet been identified. Patients with JATD have a small rib cage and narrow chest, which obstructs the lungs and can cause death by asphyxiation in infancy. In addition, the long bones, such as the femur, are short, as are the bones of the hands and feet. Patients with EVC also have short limbs and short ribs, and have dental, ectodermal, and craniofacial abnormalities.
Ruiz-Perez et al.78 showed that Indian hedgehog (Ihh) was expressed normally in the growth plates of developing bones, where it signals to induce prechondrocyte proliferation.78 However, Patched1 and Gli1 were downregulated in the bones. Similarly, Beales et al.3 showed that these target genes were downregulated in zebrafish morphants for ift80, a gene mutated in JATD.
The importance of cilia in Ihh signaling during bone development was confirmed by Koyama et al.,79 who conditionally ablated Kif3a expression in cartilage. They showed mutant growth plates lacked proliferating cells seen in controls, and lacked expression of important downstream genes such as collagen X, VEGF, and matrix metalloproteinase. Haycraft et al.80 ablated Ift88 in the limb mesenchyme, which resulted in polydactyly and shortened limbs. They showed that the polydactyly was due to Shh pathway disruption, whereas Ihh affected the extent of limb outgrowth. These data conclude that IFT is essential for normal skeletal development. The fact that some ciliopathies affect bone growth more than others might reflect different expression patterns of these genes.
We might therefore predict that cranioectodermal dysplasia (also known as Sensenbrenner syndrome), a recessive disorder similar to EVC but with the addition of renal cysts is likely caused by mutations in one of these related centrosomal/IFT genes.
Hirschsprung disease
Hirschsprung disease (HSCR) has been reported in up to 45% of patients with BBS,81 but a more realistic estimate of prevalence is 5–10% (unpublished observations of P. Beales). This incidence is extremely high compared with the general population, where it only affects 1 in 5000 children (0.02%).
HSCR arises when neurons fail to colonize the intestine, usually the colon, resulting in a lack of peristaltic activity and blockage of the gut, leading to chronic constipation and megacolon at birth and during infancy. The only treatment is surgical resection of the aganglionic segment or colostomy. Stem cell therapies to reintroduce neurons into the gut are underway in mouse models, but their efficacy is uncertain and translation to humans is a long way off.82
Tobin et al.46 showed that zebrafish deficient in Bbs8 had aganglionic guts, which were also immotile. They traced the defect to a failure of vagal neural crest cells (NCCs) to migrate into the gut. This likely explains the association of BBS with HSCR.
Craniofacial dysmorphology
Craniofacial dysmorphology is variable and can manifest as obvious and extreme, or subtle and hard to distinguish. OFD1 is an X-linked dominant lethal ciliopathy, thus all surviving patients are heterozygous females. Patients present with severe craniofacial involvement, including cleft palate and a shortened, hypoplastic midfacial region. Ferrante et al.34 produced an Ofd1 mouse knock-out with severe cleft-palate and a disorganized brain, in addition to polydactyly, renal cysts, and randomized laterality.
Abnormal craniofacial features have been reported in patients with BBS by several independent investigators, and anecdotally by family members.67,83 In contrast to OFD1, the features are variable and subtle, and it often takes a trained dysmorphologist to accurately describe the face, even though there is often an obvious gestalt, or overall appearance. Beales et al.70 described the facies of patients with BBS as having: “deep-set eyes, hypertelorism, downward slanting palpebral fissures, a flat nasal bridge with anteverted nares … a long philtrum and thin upper lip.” Lorda-Sanchez et al.83 came to similar conclusions with their analysis of the BBS face and added small mouth, malar hypoplasia (small cheek bones) and retrognathia to the facial characteristics. Figure 3 shows the typical facial appearance of children with BBS.

Facial appearance of Bardet-Biedl syndrome in children and adults. Although often subtle, characteristic features include deep-set eyes, hypertelorism, downward slanting palpebral fissures, a flat nasal bridge with anteverted nares, long philtrum and thin upper lip, small mouth, malar hypoplasia (small cheek bones), and retrognathia (small mandible).
Tobin et al.46 performed three-dimensional dense surface modeling to produce a model representing the mean features of a BBS face relative to controls. They showed that BBS faces were flattened in the mid-face region, with a hypoplastic nasal bridge and a small mandible and maxilla. This was recapitulated in both mouse and zebrafish BBS models. Real-time analysis of NCC migration in the crania of developing zebrafish embryos attributed the defect to aberrant NCC migration. Data suggested that this was due to the inhibition of the PCP pathway by knockdown of bbs8. Migrating cells had a disordered actin cytoskeleton, a feature regulated by PCP signaling. As such, it is possible that cell migration defects are present in other ciliopathies.
CILIOPATHY GENETICS AND GENOTYPE-PHENOTYPE CORRELATIONS
The first ciliopathy gene, MKKS or BBS6, was identified simultaneously by two independent groups in 2000.84,85 Since then, over 30 genes causing ciliopathy syndromes have been identified. For some of these proteins, their function is being elucidated, whereas others remain elusive. Table 2 summarizes the known ciliopathy genes and their protein products.
Most ciliopathies are autosomal recessive, with the exception of OFD1, which is X-linked dominant. However, the inheritance pattern of some ciliopathies is more complex and sometimes involves more than one locus. In 2001, it was suggested that BBS requires triallelic inheritance, consisting of homozygous mutations at one locus and a heterozygous change in a third allele at a different locus.86 Evidence for this came from homozygous mutations in BBS2 not causing disease in one individual, but when present with a third mutation in a BBS6 allele, BBS features manifested.86
Since then, it has been reported that BBS4 and BBS1 may display triallelic inheritance.87 Hoefele et al.88 screened patients with NPHP and found third alleles mutated in second NPHP loci, adding evidence that oligogenic, or triallelic, inheritance exists in other ciliopathies. Meanwhile, several studies have performed mutation screening on large cohorts of patients with BBS and found no evidence for triallelism.89–92
Leitch et al.93 found MKS1, MKS3, and CEP290 (NPHP6) mutations in patients with BBS with mutations in BBS genes. The presentation of seizures in these patients suggested that this extramutational load modified the phenotype to make it more severe. Furthermore, mutations in ciliopathy genes have been shown to cause diseases not previously associated with that gene. For example, mutations in CEP290 can cause NPHP, SLNS, or JBTS94,95; RPGRIP1L can cause MKS or JBTS,96 and AHI1 mutations can cause NPHP or JBTS.97 These findings suggest that the ciliopathies exist as a phenotypic spectrum modulated by the type, number, and position of underlying mutations. Understanding these genotype-phenotype correlations is of importance in understanding the diseases.
Furthermore, our thinking that all ciliopathies are caused by mutations in ciliary genes is being challenged. In 2008, Attanasio et al.98 identified mutations in the transcription factor gene GLIS2 in patients with NPHP and recapitulated the phenotype in mouse. Ciliary proteins have also been localized to the Golgi apparatus and to microtubules outside of the cilium99 (H. May-Simera, unpublished data). Our study of ciliopathies will need to extend beyond the cilium and basal body to fully understand the mechanisms of disease.
CILIOPATHY PROTEINS AND FUNCTIONS
Many ciliopathy proteins localize exclusively to the primary cilium (including the basal body) and/or the centriole, and so it is assumed that they function in ciliary processes. However, the primary cilium is complex and is the location of many cellular processes. These include, among others: loading and unloading of cargo on to IFT particles at the basal body, anterograde and retrograde transport along the axoneme, ciliogenesis, localizing receptors and signaling molecules to the cilium, function of signaling pathways reliant on the cilium, positioning of the cilium on the cell surface, and control of the cell cycle. Therefore, although ciliary localization is a good clue as to ciliary function, it does not delineate what that function is. This is harder to establish and so the precise function of many ciliopathy proteins at the molecular level remains unknown.
Many ciliary genes have an X-box motif in their promoter sequence to which binds the transcription factor RFX3, which activates the transcription of many ciliary genes.100 In addition, several ciliopathy proteins contain domains that give clues about their function by homology to other proteins. A few examples are given below.
MKS1
It contains a B9 domain of unknown function. In C. elegans there are three B9-containing proteins with mammalian homologues: MKS-1, TZA-1, and TZA-2.101 These form a trimeric complex at the cilium and only affect cilium formation when combined with mutations in nphp1 or nphp4. As such, the B9 domain may be effective bait for identifying new candidate ciliopathy proteins.
MKS3
Also known as Meckelin, this protein contains seven transmembrane domains, a signal peptide, four N-linked glycosylation sites, and two cysteine-rich repeats.102 This protein is structurally similar to the Frizzled receptor involved in PCP signaling. It is not yet proven, but MKS3 could play an essential, direct role in transduction of the noncanonical Wnt signal.
BBS4 and BBS8
These proteins both contain tetratricopeptide repeats, and the alternative name for these proteins are TTC10 and TTC8, respectively.5,103 Tetratricopeptide repeats are 34 amino acid motifs involved in protein-protein interactions. In addition, BBS8 contains a predicted pilF domain present in proteins involved in bacterial flagellar motility and pilus formation. This was used by Ansley et al.5 to predict its role in cilia, flagellar, or pseudopodia function.
BBS6, BBS10, and BBS12
These proteins make up a novel family of Type II chaperonin proteins involved in protein folding. Unlike any other BBS genes, they are specific to vertebrates, rapidly evolving, and may function redundantly.104,105 It is unknown what the function of these chaperonins is, but they are not included in the interacting complex of BBS proteins, the BBSome (see Nachury et al.106), but may be involved in aiding the folding of other BBS or IFT proteins.
Functional studies reveal that the protein products of MKS1 and MKS3/TMEM67 (meckelin) interact with each other and are required for centriole migration to the apical membrane and consequent formation of the primary cilium.19 Recently, a sixth locus for MKS was identified and the gene MKS6/CC2D2A reported; although the biological function of CC2D2A is unknown, the identification of a calcium binding domain and the fact that patient-derived fibroblasts lack cilia indicate a critical role for CC2D2A in cilia function.126,127
The AHI1 gene product is Jouberin. It is expressed in the brain and kidney and has been shown to interact with nephrocystin-1. In a mouse kidney cell line, AHI1 was shown to localize at adherens junctions, the primary cilia and basal body, which is consistent with a role in NPHP.128
No structural data from crystallographic studies has been obtained, largely owing to the fact that BBS proteins are insoluble. However, function is better characterized for BBS4, BBS7, BBS8, IFT80, and NEK8, from which inferences about other ciliopathy protein functions have been made. We outline four studies that have illuminated functions of the BBS proteins.
The first clues about function came from studies of BBS-7 and BBS-8 in C. elegans. Blacque et al.117 first showed that these two proteins localize to the base of the cilium in ciliated neurons, and GFP-tagged proteins could be observed participating in IFT. The average rate of anterograde movement was approximately 0.7 μm s−1, whereas retrograde was roughly 1.1 μm s−1; rates comparable with IFT motor proteins such as OSM-3 kinesin. The authors found animals mutant for these two genes were defective in taxis toward two chemical attractants whose detection relies upon ciliated neurons. They then showed that cilia structure was abnormal in these mutants, determined by the inability to uptake DiI. GFP-tagged IFT proteins failed to localize and move properly along the cilium in bbs-7 and bbs-8 mutants. The authors suggested that these proteins facilitate the incorporation of the IFT particle onto the motor protein complex, and are selective for particular cargoes.
Ou et al.129 used the same C. elegans mutants to show that BBS-7 and BBS-8 coordinate the movement of IFT subcomplexes A and B. In the absence of these proteins, the subcomplexes moved separately, at different rates, because a different type of kinesin motor moves each subcomplex. The authors proposed a model whereby BBS-7 and BBS-8 act as a bridge between the two IFT subcomplexes. In the absence of these proteins, the two subcomplexes move independently and are unstable. As such, IFT is not completely abolished in bbs mutants, but it becomes unstable and slow. This explains why BBS phenotypes are less severe than complete IFT knockouts, and are not lethal.
Kim et al.130 performed a yeast two-hybrid screen to identify p150 and PCM1 as interactors of BBS4. Silencing of Bbs4 resulted in abrogation of recruitment of PCM1 to the pericentriolar satellites, and a deanchoring of microtubules from the centrosome indicating BBS4 behaves as an adaptor protein.
Nachury et al.106 also provided insight as to how BBS proteins behave at the cellular level. They used tandem affinity purification to isolate a complex of BBS proteins they termed the “BBSome.” This 438kDa complex consisted of stoichiometric ratios of BBS1, 2, 4, 5, 7, 8, and 9, with BBS9 acting as the central organizer, as it interacted with all other subunits. They found the most marked defects of ciliogenesis when cells were depleted of BBS1 and 5. They conjectured that the BBSome is transported to the basal body by the centriolar satellites and it associates with the ciliary membrane. They showed the BBSome to interact with Rabin8, a guanosyl exchange factor for Rab8, a small GTPase that regulates vesicle trafficking from post-Golgi vesicles. Rab8 enters the ciliary membrane and promotes ciliogenesis. They proposed that pathogenesis of BBS might be caused by defects in vesicular transport to the cilium.
It was recently shown that CEP290, whose gene is mutated in MKS and JBS, interacts with PCM1, as BBS4 does.131 Furthermore, CEP290 is required for localization of Rab8 to the primary cilium, in keeping with data on the BBSome. The authors suggested that PCM1 might link CEP290 to BBS proteins in common molecular pathways. It is possible that there are many more interacting partners among the ciliopathy proteins. Future proteomic analyses may reveal a “ciliopathome,” in which many ciliopathy proteins interact, explaining their convergent phenotypes, despite often having no sequence homology.
In light of these studies, it seems there are two general, nonexclusive models for how BBS proteins might function. One suggests that at least BBS4, 7, and 8 are necessary for microtubule-based transport along the cilium. The other theory implies that BBS proteins function as a complex mediating vesicle transport from the cytoplasm to the cilium. As such, no one clear model for the function of BBS proteins has been described and the proteins may function in numerous roles, including cytoplasmic transport.
MOUSE MODELS OF CILIOPATHIES
Mouse models exist for BBS (Bbs1, Bbs1 M390R knock-in, Bbs2, Bbs4, and Bbs6); ALMS (Alms1 or fat aussie), NPHP (Nphp1, Nphp2, Nphp3, Cep290/Nphp6); ofd1; and EVC (Evc1). Here, some of these models will be discussed, highlighting their contribution to understanding the disease.
Bbs1 knockout
This model was first reported by Kulaga et al.132 in 2004. They showed that BBS1 and Bbs4 nulls, had absent cilia on the olfactory, but not the respiratory, epithelia. Adult mice were anosmic, a finding that was extended to humans to reveal a previously unrecognized clinical feature of the disease.
This mouse model was used in a subsequent study by Tan et al.133 to show that mouse mutants for Bbs1 and Bbs4 had impaired sensory perception of heat and pain. Again, this was used to extend studies to patients, who had reduced response to painful stimuli. These defects were attributed to aberrant localization of thermosensory and mechanosensory ion channels in the receptors.
Bbs1 M390R knock-in
Davis et al.134 published a model of BBS where they used homologous recombination to create a transgenic mouse expressing the commonest point mutation in white patients with BBS, M390R. They showed that these mice had all the cardinal features of BBS, including retinal degeneration, obesity, and infertility. They also found swollen brain ventricles, reduced size of the striatum and hippocampus, and a thin cerebral cortex.
Bbs2 knockout
Nishimura et al.135 produced a knockout of Bbs2. These animals recapitulated the common BBS phenotypes such as obesity and retinal degeneration, but in keeping with other mouse models, were not polydactylous. This model revealed that the mutants lacked social dominance and were always outcompeted in tests against heterozygous littermates. In addition, clues about retinal degeneration were gathered from the mislocalization of rhodopsin in the rod cells, and increased apoptotic cell death of rods.
Another study showed that these mutants, as well as Bbs4 and Bbs6 nulls, had elevated levels of serum leptin and they suggested that these animals were leptin resistant. They also found decreased expression of proopiomelanocortin in the hypothalamus. They implied that BBS genes were important for appetite regulation.
Bbs4 knockout
This mutant was first published by Mykytyn et al.110 who showed that, in addition to the above mentioned phenotypes, these mutants' sperm lacked flagella, and as such, nulls were infertile. This also raises the possibility that BBS proteins, as well as other nonmotile ciliopathy proteins, may be involved in the construction and maintenance of motile cilia as well.
Ross et al.49 used Bbs4 null mice to implicate the genes in the PCP pathway. A proportion (14%) of Bbs4 null embryos had PCP-like phenotypes, such as open eyelids and exencephaly. NTDs (e.g., encephalocoele) are also prevalent in other ciliopathies such as MKS and JBSs. Disordered stereociliary bundles of hair cells in the inner ear led to clinical audiological testing, which revealed subclinical hearing loss in patients with BBS. This is another instance of where mouse models have informed clinical practice.
Eichers et al.136 performed a more detailed analysis of the Bbs4 null mouse and found some new phenotypes. They found that females suffered more severe obesity than their male counterparts did, and that blood chemistry showed lipid profiles similar to the metabolic syndrome. This was indicative of liver dysfunction, something not well investigated in BBS. They also noted the rare existence of NTDs, as well as genital abnormalities, such as hydrometrocolpos.
EVC knockout
Evc−/− mice develop an EVC-like syndrome, including short ribs, short limbs, and dental abnormalities.78 Evc is expressed in the developing bones and the orofacial region and at the cellular level Evc is located at the base of the primary cilium. Although, Ihh is expressed normally in the growth plates of Evc−/− mice, expression of Ihh downstream genes Ptch1 and Gli1 was markedly decreased.
Mks1 knockout
A mouse mutant named kerouac (krc) was recently identified from an ethyl-N-nitrosourea mutagenesis screen.137 Its phenotype included preaxial polydactyly, bowing of the long bones of the limbs, left-right asymmetry defects, polycystic kidneys, defects in the ribs and sternum, ventral patterning defects in the neural tube and perinatal lethality. Krc maps to the Mks1 gene locus therefore explaining the partial MKS phenotype recapitulation.
Nphp2 knockout
The inv mutation in mouse was originally created by a random insertional mutagenesis and subsequently shown to encode the gene Inversin.138,139 Otto et al. then found that the human orthologue of Inversin was mutated in patients with NPHP. They named this gene NPHP2. This provided a clear link between a gene that was known to function in microtubule-associated processes at the node of the embryo, with laterality defects in mice and humans.
Nphp3 knockout
Bergmann et al.57 showed that the common mouse mutant pcy (polycystic kidneys) contained hypomorphic mutations in Nphp3, a gene mutated in human NPHP. They also showed that null mutations in Nphp3 were embryonic lethal in mice, whereas hypomorphs survived. Nphp3 interacts with Inversin, and like Inversin, might act as a switch to control the canonical versus noncanonical Wnt signaling pathways.
Ftm knockout
The mouse model of Ftm (Rpgrip1l) was published before it was identified as a causative gene of MKS and JBTS.45,96 The Ftm null mouse had aberrant Shh signaling; including altered processing of the transcription factor Gli3. Because Ftm was not required for ciliary assembly, they suggested that Ftm was a novel ciliary protein required for Shh signaling. This could apply to other ciliopathy proteins, particularly in light of the fact that Shh transduction defects have been observed when Bbs8 is repressed.46
Ofd1 knockout
This mouse mutant is unusual because OFD1 is an X-linked male lethal disease; so all mice born are female. Ferrante et al.34 showed that heterozygous female mice had cystic kidneys which lacked primary cilia, had situs inversus, and polydactyly. They also showed misexpression of Hoxa cluster genes, suggesting that Ofd1 has a role in fundamental embryo organization.
This mutant was used in another study to show that these embryos, which lack primary cilia, had elevated canonical Wnt signaling.53 Combining these data with two other mouse mutants (Kif3a and Ift88), these authors suggested that the primary cilium acts as an inhibitor of the canonical Wnt pathway, and that abrogation of it upregulates canonical Wnt signaling.
It can be seen from the above examples how useful the mouse models have been, both for better understanding of the human disease, and for giving insights into the basic biology that ciliopathy proteins are important for.
THERAPIES FOR CILIOPATHIES
Development of orphan drugs to treat diseases as rare as the ciliopathies is economically unfeasible. As the ciliopathies present with complex, multisystem involvement, the best hope for treatment is to ameliorate the features of the disease, which cause the greatest burden to health. In the case of BBS, this includes renal disease, blindness, and obesity. As MKS and JBTS are both lethal, the focus should be on providing better prenatal diagnosis.
Kidney failure causes the greatest degradation to quality of life of the ciliopathy features and is the most widespread among the diseases. Because the features of the kidney pathology are similar to common genetic forms of PKD, it may be possible that candidate molecules used to treat these diseases can be trialed on patients with ciliopathy. A better understanding of the molecular and cellular etiology of kidney cystogenesis and dysplasia in diseases such as BBS and NPHP, combined with mouse models, would enable preclinical trials to take place.
ADPKD is one of the most common genetic diseases, with an incidence of around 1 in 1000. The onset of the disease is much later than autosomal recessive polycystic kidney disease (ARPKD), with a median of around 15 years.140 Cysts grow progressively larger and kidney mass has been reported to reach up to 20 kg.141 As such, therapies to slow the development of the cysts prolong symptom-free status, as the health only becomes compromised when the cysts impair the filtering capacity of the kidney. This results in ESRF in approximately 50% of patients with ADPKD.
Currently there is no treatment for PKD, there are only drugs that slow cyst development, with some promising molecules in preclinical development in the mouse, but no compounds have entered the market to treat humans yet. Present treatment involves a low-protein diet to offset renal failure. Current advances in understanding the mechanism of cystogenesis will inform drug discovery strategies. Several important mouse models are being used as PKD models: the orpk mouse (Oak Ridge PKD) is a polaris mutant, which encodes IFT88; the inv mouse in which Nphp2 is mutated; and conditional knock-out of Kif3a in the kidney causes cysts.
Current pathways targeted for therapy include the Ras pathway, a mitogenic signaling pathway upregulated in PKD. Ras signals via Raf to translocate the MAP kinase ERK to the nucleus, whereupon it phosphorylates transcription factors associated with cell division. Another pathway involved in cyst progression is the GPCR pathway, where cAMP activates Raf kinase activity to drive cell proliferation. Therefore, cAMP inhibitors are another option for therapy.
Using zebrafish to screen molecules
Tobin and Beales142 provided the first data to suggest that the zebrafish could be used to quantitatively assess the therapeutic potential of candidate molecules for ciliopathic renal disease. By generating knockdown models of 10 different ciliopathy genes, they showed that Rapamycin and Roscovitine were able to ameliorate both the formation of a renal cyst, as well as the filtering capacity of the kidney. Rapamycin and Roscovitine have both been previously shown to be effective at treating mouse models of PKD.55,143
The ease with which genes can be knocked down in the zebrafish, the huge clutch sizes, the speed of cyst development, the ease of scoring the phenotype, the physiological relevance, and simplicity of adding drugs, and the scalability, make the zebrafish an ideal model for testing therapeutics for rare diseases. These then inform a more directed strategy to test compounds on mouse models.
Strategies to identify new ciliopathies
Beales et al.3 showed that the core features of ciliopathies are so consistent and reproducible, they can be used to predict a disease with ciliary involvement. Looking at current diseases, it is possible that several genes not currently classified as ciliopathies, may, in the future, be proven to be so. Carpenter syndrome involves polydactyly, craniofacial dysmorphology, obesity, mental retardation, and hypogonadism. The gene mutated in Carpenter syndrome is RAB23. This gene product is a negative regulator of Shh signaling involved in vesicle transport.144 Rab23 knockout mice have phenotypes reminiscent of other ciliary Shh knockouts, such as Ift172 and Ift88. Huangfu et al.40 showed that Ift proteins were required for the activity of Rab23.
These data provide compelling evidence that Carpenter syndrome may be a novel ciliopathy; however, definitive proof will require the ciliary localization of Rab23. However, it is possible that not all ciliopathy proteins localize to the cilium. Pathways such as Shh and Wnt signal to the nucleus via the cilium. As such, the signal is transduced from the cilium, via the cytoplasm, to the nucleus. It is possible that some of the ciliopathy proteins intervene at the “postcilium” stages of signaling, either en route to the nucleus or in the nucleus itself. This may be particularly true for trafficking proteins like Rab23, and is pertinent in light of the fact that Rab8 interacts with the BBSome, and knockdown in the zebrafish recapitulates BBS phenotypes.
Another putative ciliopathy is Acrocallosal syndrome comprising agenesis of the corpus callosum, postaxial-polydactyly, and mental retardation. Some patients have been ascribed mutations in GLI3, but these do not account for all mutations. It is probable that mutations in ciliary genes account for some cases.
As it becomes clearer the precise cellular events and signaling pathways in which the cilium partakes, it will be easier to attribute ciliopathies. The accessibility of high-throughput sequencing makes it easy to screen large sets of ciliary genes in large patient cohorts to identify mutations in novel disease genes. Given the size of the ciliary proteome (1000 proteins), it is likely that many more of these genes will harbor disease mutations. The future of ciliopathy genetics will involve medical reseqeuncing of dozens of genes in large patient cohorts to identify new causative genes and define genotype-phenotype relationships.
Future areas of study
As our knowledge of the ciliopathies increases from cell biological, human genetic, and animal model studies, we still know very little about the function of ciliopathy proteins at the molecular and atomic levels. As such, the next critical step in understanding ciliary biology will be to undertake structural biology studies to determine crystal structures of the IFT complexes, the ciliome, and other key ciliopathy proteins.
In addition, we have very poor understanding of how the few ciliopathies from known protein families, such as the chaperonins (BBS6, 10, and 12), and the E3 ubiquitin ligase (BBS11), function. Biochemical assays to determine what the potential substrates of these molecules are, if any, will be important to understand their function.
As the number and expanse of ciliopathies and associated genes increases, it will become increasingly important to do genome, or ciliome-wide screens to identify novel interactions of these diverse proteins. A comprehensive ciliopathy interactome will provide a huge resource for understanding the disease and beginning to piece together genotype-phenotype correlations that might be useful to make clinical predictions.
Alongside these functional studies, large-scale gene sequencing efforts to resequence all the genes of the ciliary proteome will establish how these mutations give rise to the human phenotypes. This has been started by Leitch et al.,90 who showed that mutations previously thought to be responsible for other ciliopathies are found in BBS, and that they are able to modify the phenotype. The ever-reducing cost of large-scale sequencing will enable the identification of mutations in novel genes that produce or modify the ciliopathy phenotype.
References
Yoder BK . Role of primary cilia in the pathogenesis of polycystic kidney disease. J Am Soc Nephrol 2007; 18: 1381–1388.
Simons M, Walz G . Polycystic kidney disease: cell division without a c(l)ue?. Kidney Int 2006; 70: 854–864.
Beales PL, Bland E, Tobin JL, et al. IFT80, which encodes a conserved intraflagellar transport protein, is mutated in Jeune asphyxiating thoracic dystrophy. Nat Genet 2007; 39: 727–729.
Hildebrandt F, Zhou W . Nephronophthisis-associated ciliopathies. J Am Soc Nephrol 2007; 18: 1855–1871.
Ansley SJ, Badano JL, Blacque OE, et al. Basal body dysfunction is a likely cause of pleiotropic Bardet-Biedl syndrome. Nature 2003; 425: 628–633.
Berbari NF, Johnson AD, Lewis JS, Askwith CC, Mykytyn K . Identification of ciliary localization sequences within the third intracellular loop of G protein-coupled receptors. Mol Biol Cell 2008; 19: 1540–1547.
Berbari NF, Lewis JS, Bishop GA, Askwith CC, Mykytyn K . Bardet-Biedl syndrome proteins are required for the localization of G protein-coupled receptors to primary cilia. Proc Natl Acad Sci USA 2008; 105: 4242–4246.
Alton DJ, McDonald P . Urographic findings in the Bardet-Biedl syndrome, formerly the Laurence-Moon-Biedl syndrome. Radiology 1973; 109: 659–663.
Joy T, Cao H, Black G, et al. Alstrom syndrome (OMIM 203800): a case report and literature review. Orphanet J Rare Dis 2007; 2: 49.
Collin GB, Marshall JD, Ikeda A, et al. Mutations in ALMS1 cause obesity, type 2 diabetes and neurosensory degeneration in Alstrom syndrome. Nat Genet 2002; 31: 74–78.
Hearn T, Renforth GL, Spalluto C, et al. Mutation of ALMS1, a large gene with a tandem repeat encoding 47 amino acids, causes Alstrom syndrome. Nat Genet 2002; 31: 79–83.
Andersen JS, Wilkinson CJ, Mayor T, Mortensen P, Nigg EA, Mann M . Proteomic characterization of the human centrosome by protein correlation profiling. Nature 2003; 426: 570–574.
Hearn T, Spalluto C, Phillips VJ, et al. Subcellular localization of ALMS1 supports involvement of centrosome and basal body dysfunction in the pathogenesis of obesity, insulin resistance, and type 2 diabetes. Diabetes 2005; 54: 1581–1587.
Collin GB, Cyr E, Bronson R, et al. Alms1-disrupted mice recapitulate human Alstrom syndrome. Hum Mol Genet 2005; 14: 2323–2333.
Arsov T, Silva DG, O'Bryan MK, et al. Fat aussie—a new Alstrom syndrome mouse showing a critical role for ALMS1 in obesity, diabetes, and spermatogenesis. Mol Endocrinol 2006; 20: 1610–1622.
Salonen R . The Meckel syndrome: clinicopathological findings in 67 patients. Am J Med Genet 1984; 18: 671–689.
Salonen R, Paavola P . Meckel syndrome. J Med Genet 1998; 35: 497–501.
Paavola P, Salonen R, Baumer A, et al. Clinical and genetic heterogeneity in Meckel syndrome. Hum Genet 1997; 101: 88–92.
Dawe HR, Smith UM, Cullinane AR, et al. The Meckel-Gruber Syndrome proteins MKS1 and meckelin interact and are required for primary cilium formation. Hum Mol Genet 2007; 16: 173–186.
Louie CM, Gleeson JG . Genetic basis of Joubert syndrome and related disorders of cerebellar development. Hum Mol Genet 2005; 14: R235–R242.
Valente EM, Salpietro DC, Brancati F, et al. Description, nomenclature, and mapping of a novel cerebello-renal syndrome with the molar tooth malformation. Am J Hum Genet 2003; 73: 663–670.
Dixon-Salazar T, Silhavy JL, Marsh SE, et al. Mutations in the AHI1 gene, encoding jouberin, cause Joubert syndrome with cortical polymicrogyria. Am J Hum Genet 2004; 75: 979–987.
Ferland RJ, Eyaid W, Collura RV, et al. Abnormal cerebellar development and axonal decussation due to mutations in AHI1 in Joubert syndrome. Nat Genet 2004; 36: 1008–1013.
Valente EM, Brancati F, Silhavy JL, et al. AHI1 gene mutations cause specific forms of Joubert syndrome-related disorders. Ann Neurol 2006; 59: 527–534.
Utsch B, Sayer JA, Attanasio M, et al. Identification of the first AHI1 gene mutations in nephronophthisis-associated Joubert syndrome. Pediatr Nephrol 2006; 21: 32–35.
Parisi MA, Doherty D, Eckert ML, et al. AHI1 mutations cause both retinal dystrophy and renal cystic disease in Joubert syndrome. J Med Genet 2006; 43: 334–339.
Cantagrel V, Silhavy JL, Bielas SL, et al. Mutations in the cilia gene ARL13B lead to the classical form of Joubert syndrome. Am J Hum Genet 2008; 83: 170–179.
Garcia-Garcia MJ, Eggenschwiler JT, Caspary T, et al. Analysis of mouse embryonic patterning and morphogenesis by forward genetics. Proc Natl Acad Sci USA 2005; 102: 5913–5919.
Sun Z, Amsterdam A, Pazour GJ, Cole DG, Miller MS, Hopkins N . A genetic screen in zebrafish identifies cilia genes as a principal cause of cystic kidney. Development 2004; 131: 4085–4093.
Caspary T, Larkins CE, Anderson KV . The graded response to Sonic Hedgehog depends on cilia architecture. Dev Cell 2007; 12: 767–778.
Baujat G, Le Merrer M . Ellis-van Creveld syndrome. Orphanet J Rare Dis 2007; 2: 27.
Ferrante MI, Giorgio G, Feather SA, et al. Identification of the gene for oral-facial-digital type I syndrome. Am J Hum Genet 2001; 68: 569–576.
Giorgio G, Alfieri M, Prattichizzo C, Zullo A, Cairo S, Franco B . Functional characterization of the OFD1 protein reveals a nuclear localization and physical interaction with subunits of a chromatin remodeling complex. Mol Biol Cell 2007; 18: 4397–4404.
Ferrante MI, Zullo A, Barra A, et al. Oral-facial-digital type I protein is required for primary cilia formation and left-right axis specification. Nat Genet 2006; 38: 112–117.
Webber WA, Lee J . Fine structure of mammalian renal cilia. Anat Rec 1975; 182: 339–343.
Rosenbaum JL, Witman GB . Intraflagellar transport. Nat Rev Mol Cell Biol 2002; 3: 813–825.
Lehman JM, Michaud EJ, Schoeb TR, Aydin-Son Y, Miller M, Yoder BK . The Oak Ridge Polycystic Kidney mouse: modeling ciliopathies of mice and men. Dev Dyn 2008; 237: 1960–1971.
Gherman A, Davis EE, Katsanis N . The ciliary proteome database: an integrated community resource for the genetic and functional dissection of cilia. Nat Genet 2006; 38: 961–962.
Davis EE, Brueckner M, Katsanis N . The emerging complexity of the vertebrate cilium: new functional roles for an ancient organelle. Dev Cell 2006; 11: 9–19.
Huangfu D, Liu A, Rakeman AS, Murcia NS, Niswander L, Anderson KV . Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature 2003; 426: 83–87.
Liu A, Wang B, Niswander LA . Mouse intraflagellar transport proteins regulate both the activator and repressor functions of Gli transcription factors. Development 2005; 132: 3103–3111.
Corbit KC, Aanstad P, Singla V, Norman AR, Stainier DY, Reiter JF . Vertebrate Smoothened functions at the primary cilium. Nature 2005; 437: 1018–1021.
Rohatgi R, Milenkovic L, Scott MP . Patched1 regulates hedgehog signaling at the primary cilium. Science 2007; 317: 372–376.
May SR, Ashique AM, Karlen M, et al. Loss of the retrograde motor for IFT disrupts localization of Smo to cilia and prevents the expression of both activator and repressor functions of Gli. Dev Biol 2005; 287: 378–389.
Vierkotten J, Dildrop R, Peters T, Wang B, Rüther U . Ftm is a novel basal body protein of cilia involved in Shh signalling. Development 2007; 134: 2569–2577.
Tobin JL, Di Franco M, Eichers E, et al. Inhibition of neural crest migration underlies craniofacial dysmorphology and Hirschsprung's disease in Bardet-Biedl syndrome. Proc Natl Acad Sci USA 2008; 105: 6714–6719.
Fischer E, Legue E, Doyen A, et al. Defective planar cell polarity in polycystic kidney disease. Nat Genet 2006; 38: 21–23.
Simons M, Gloy J, Ganner A, et al. Inversin, the gene product mutated in nephronophthisis type II, functions as a molecular switch between Wnt signaling pathways. Nat Genet 2005; 37: 537–543.
Ross AJ, May-Simera H, Eichers ER, et al. Disruption of Bardet-Biedl syndrome ciliary proteins perturbs planar cell polarity in vertebrates. Nat Genet 2005; 37: 1135–1140.
Park TJ, Haigo SL, Wallingford JB . Ciliogenesis defects in embryos lacking inturned or fuzzy function are associated with failure of planar cell polarity and Hedgehog signaling. Nat Genet 2006; 38: 303–311.
Park TJ, Mitchell BJ, Abitua PB, Kintner C, Wallingford JB . Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells. Nat Genet 2008; 40: 871–879.
Gerdes JM, Liu Y, Zaghloul NA, et al. Disruption of the basal body compromises proteasomal function and perturbs intracellular Wnt response. Nat Genet 2007; 39: 1350–1360.
Corbit KC, Shyer AE, Dowdle WE, et al. Kif3a constrains beta-catenin-dependent Wnt signalling through dual ciliary and non-ciliary mechanisms. Nat Cell Biol 2008; 10: 70–76.
Edgar BA . How flies get their size: genetics meets physiology. Nat Rev Genet 2006; 7: 907–916.
Shillingford JM, Murcia NS, Larson CH, et al. The mTOR pathway is regulated by polycystin-1, and its inhibition reverses renal cystogenesis in polycystic kidney disease. Proc Natl Acad Sci USA 2006; 103: 5466–5471.
Dibella LM, Park A, Sun Z . Zebrafish Tsc1 reveals functional interactions between the cilium and the TOR pathway. Hum Mol Genet 2008; 18: 595–606.
Bergmann C, Fliegauf M, Brüchle NO, et al. Loss of nephrocystin-3 function can cause embryonic lethality, Meckel-Gruber-like syndrome, situs inversus, and renal-hepatic-pancreatic dysplasia. Am J Hum Genet 2008; 82: 959–970.
Klysik M . Ciliary syndromes and treatment. Pathol Res Pract 2008; 204: 77–88.
Lorda-Sanchez I, Ayuso C, Ibañez A . Situs inversus and hirschsprung disease: two uncommon manifestations in Bardet-Biedl syndrome. Am J Med Genet 2000; 90: 80–81.
Otto EA, Schermer B, Obara T, et al. Mutations in INVS encoding inversin cause nephronophthisis type 2, linking renal cystic disease to the function of primary cilia and left-right axis determination. Nat Genet 2003; 34: 413–420.
Brueckner M, McGrath J, D'Eustachio P, Horwich AL . Establishment of left-right asymmetry in vertebrates: genetically distinct steps are involved. Ciba Found Symp 1991; 162: 202–212; discussion 212–208.
McGrath J, Brueckner M . Cilia are at the heart of vertebrate left-right asymmetry. Curr Opin Genet Dev 2003; 13: 385–392.
Tanaka Y, Okada Y, Hirokawa N . FGF-induced vesicular release of Sonic hedgehog and retinoic acid in leftward nodal flow is critical for left-right determination. Nature 2005; 435: 172–177.
McGrath J, Somlo S, Makova S, Tian X, Brueckner M . Two populations of node monocilia initiate left-right asymmetry in the mouse. Cell 2003; 114: 61–73.
Watanabe D, Saijoh Y, Nonaka S, et al. The left-right determinant Inversin is a component of node monocilia and other 9+0 cilia. Development 2003; 130: 1725–1734.
Oishi I, Kawakami Y, Raya A, Callol-Massot C, Izpisúa Belmonte JC . Regulation of primary cilia formation and left-right patterning in zebrafish by a noncanonical Wnt signaling mediator, duboraya. Nat Genet 2006; 38: 1316–1322.
Beales PL, Elcioglu N, Woolf AS, Parker D, Flinter FA . New criteria for improved diagnosis of Bardet-Biedl syndrome: results of a population survey. J Med Genet 1999; 36: 437–446.
Abd-El-Barr MM, Sykoudis K, Andrabi S, et al. Impaired photoreceptor protein transport and synaptic transmission in a mouse model of Bardet-Biedl syndrome. Vision Res 2007; 47: 3394–3407.
Bainbridge JW, Smith AJ, Barker SS, et al. Effect of gene therapy on visual function in Leber's congenital amaurosis. N Engl J Med 2008; 358: 2231–2239.
Marshall JD, Beck S, Maffei P, Naggert JK . Alström syndrome. Eur J Hum Genet 2007; 15: 1193–1202.
Li G, Vega R, Nelms K, et al. A role for Alström syndrome protein, alms1, in kidney ciliogenesis and cellular quiescence. PLoS Genet 2007; 3: e8.
Green JS, Parfrey PS, Harnett JD, et al. The cardinal manifestations of Bardet-Biedl syndrome, a form of Laurence-Moon-Biedl syndrome. N Engl J Med 1989; 321: 1002–1009.
Davenport JR, Watts AJ, Roper VC, et al. Disruption of intraflagellar transport in adult mice leads to obesity and slow-onset cystic kidney disease. Curr Biol 2007; 17: 1586–1594.
Grace C, Beales PL, Summerbell C, et al. Energy metabolism in Bardet-Biedl syndrome. Int J Obes Relat Metab Disord 2003; 27: 1319–1324.
Rahmouni K, Fath MA, Seo S, et al. Leptin resistance contributes to obesity and hypertension in mouse models of Bardet-Biedl syndrome. J Clin Invest 2008; 118: 1458–1467.
Romano S, Milan G, Veronese C, et al. Regulation of Alström syndrome gene expression during adipogenesis and its relationship with fat cell insulin sensitivity. Int J Mol Med 2008; 21: 731–736.
Forti E, Aksanov O, Birk RZ . Temporal expression pattern of Bardet-Biedl syndrome genes in adipogenesis. Int J Biochem Cell Biol 2007; 39: 1055–1062.
Ruiz-Perez VL, Blair HJ, Rodriguez-Andres ME, et al. Evc is a positive mediator of Ihh-regulated bone growth that localises at the base of chondrocyte cilia. Development 2007; 134: 2903–2912.
Koyama E, Young B, Nagayama M, et al. Conditional Kif3a ablation causes abnormal hedgehog signaling topography, growth plate dysfunction, and excessive bone and cartilage formation during mouse skeletogenesis. Development 2007; 134: 2159–2169.
Haycraft CJ, Zhang Q, Song B, et al. Intraflagellar transport is essential for endochondral bone formation. Development 2007; 134: 307–316.
de Pontual L, Pelet A, Clement-Ziza M, et al. Epistatic interactions with a common hypomorphic RET allele in syndromic Hirschsprung disease. Hum Mutat 2007; 28: 790–796.
Gershon MD . Transplanting the enteric nervous system: a step closer to treatment for aganglionosis. Gut 2007; 56: 459–461.
Lorda-Sanchez I, Ayuso C, Sanz R, Ibañez A . Does Bardet-Biedl syndrome have a characteristic face?. J Med Genet 2001; 38: E14.
Slavotinek AM, Stone EM, Mykytyn K, et al. Mutations in MKKS cause Bardet-Biedl syndrome. Nat Genet 2000; 26: 15–16.
Katsanis N, Beales PL, Woods MO, et al. Mutations in MKKS cause obesity, retinal dystrophy and renal malformations associated with Bardet-Biedl syndrome. Nat Genet 2000; 26: 67–70.
Katsanis N, Ansley SJ, Badano JL, et al. Triallelic inheritance in Bardet-Biedl syndrome, a Mendelian recessive disorder. Science 2001; 293: 2256–2259.
Beales PL, Badano JL, Ross AJ, et al. Genetic interaction of BBS1 mutations with alleles at other BBS loci can result in non-Mendelian Bardet-Biedl syndrome. Am J Hum Genet 2003; 72: 1187–1199.
Hoefele J, Wolf MT, O'Toole JF, et al. Evidence of oligogenic inheritance in nephronophthisis. J Am Soc Nephrol 2007; 18: 2789–2795.
Hichri H, Stoetzel C, Laurier V, et al. Testing for triallelism: analysis of six BBS genes in a Bardet-Biedl syndrome family cohort. Eur J Hum Genet 2005; 13: 607–616.
Nakane T, Biesecker LG . No evidence for triallelic inheritance of MKKS/BBS loci in Amish Mckusick-Kaufman syndrome. Am J Med Genet A 2005; 138: 32–34.
Smaoui N, Chaabouni M, Sergeev YV, et al. Screening of the eight BBS genes in Tunisian families: no evidence of triallelism. Invest Ophthalmol Vis Sci 2006; 47: 3487–3495.
Laurier V, Stoetzel C, Muller J, et al. Pitfalls of homozygosity mapping: an extended consanguineous Bardet-Biedl syndrome family with two mutant genes (BBS2, BBS10), three mutations, but no triallelism. Eur J Hum Genet 2006; 14: 1195–1203.
Leitch CC, Zaghloul NA, Davis EE, et al. Hypomorphic mutations in syndromic encephalocele genes are associated with Bardet-Biedl syndrome. Nat Genet 2008; 40: 443–448.
Baala L, Audollent S, Martinovic J, et al. Pleiotropic effects of CEP290 (NPHP6) mutations extend to Meckel syndrome. Am J Hum Genet 2007; 81: 170–179.
Helou J, Otto EA, Attanasio M, et al. Mutation analysis of NPHP6/CEP290 in patients with Joubert syndrome and Senior-Løken syndrome. J Med Genet 2007; 44: 657–663.
Delous M, Baala L, Salomon R, et al. The ciliary gene RPGRIP1L is mutated in cerebello-oculo-renal syndrome (Joubert syndrome type B) and Meckel syndrome. Nat Genet 2007; 39: 875–881.
Wolf MT, Saunier S, O'Toole JF, et al. Mutational analysis of the RPGRIP1L gene in patients with Joubert syndrome and nephronophthisis. Kidney Int 2007; 72: 1520–1526.
Attanasio M, Uhlenhaut NH, Sousa VH, et al. Loss of GLIS2 causes nephronophthisis in humans and mice by increased apoptosis and fibrosis. Nat Genet 2007; 39: 1018–1024.
Follit JA, Tuft RA, Fogarty KE, Pazour GJ . The intraflagellar transport protein IFT20 is associated with the Golgi complex and is required for cilia assembly. Mol Biol Cell 2006; 17: 3781–3792.
Efimenko E, Bubb K, Mak HY, et al. Analysis of xbx genes in C. elegans. Development 2005; 132: 1923–1934.
Williams CL, Winkelbauer ME, Schafer JC, Michaud EJ, Yoder BK . Functional redundancy of the B9 proteins and nephrocystins in Caenorhabditis elegans ciliogenesis. Mol Biol Cell 2008; 19: 2154–2168.
Smith UM, Consugar M, Tee LJ, et al. The transmembrane protein meckelin (MKS3) is mutated in Meckel-Gruber syndrome and the wpk rat. Nat Genet 2006; 38: 191–196.
Mykytyn K, Braun T, Carmi R, et al. Identification of the gene that, when mutated, causes the human obesity syndrome BBS4. Nat Genet 2001; 28: 188–191.
Stoetzel C, Muller J, Laurier V, et al. Identification of a novel BBS gene (BBS12) highlights the major role of a vertebrate-specific branch of chaperonin-related proteins in Bardet-Biedl syndrome. Am J Hum Genet 2007; 80: 1–11.
Stoetzel C, Laurier V, Davis EE, et al. BBS10 encodes a vertebrate-specific chaperonin-like protein and is a major BBS locus. Nat Genet 2006; 38: 521–524.
Nachury MV, Loktev AV, Zhang Q, et al. A core complex of BBS proteins cooperates with the GTPase Rab8 to promote ciliary membrane biogenesis. Cell 2007; 129: 1201–1213.
Mykytyn K, Nishimura DY, Searby CC, et al. Identification of the gene (BBS1) most commonly involved in Bardet-Biedl syndrome, a complex human obesity syndrome. Nat Genet 2002; 31: 435–438.
Nishimura DY, Searby CC, Carmi R, et al. Positional cloning of a novel gene on chromosome 16q causing Bardet-Biedl syndrome (BBS2). Hum Mol Genet 2001; 10: 865–874.
Fan Y, Esmail MA, Ansley SJ, et al. Mutations in a member of the Ras superfamily of small GTP-binding proteins causes Bardet-Biedl syndrome. Nat Genet 2004; 36: 989–993.
Mykytyn K, Mullins RF, Andrews M, et al. Bardet-Biedl syndrome type 4 (BBS4)-null mice implicate Bbs4 in flagella formation but not global cilia assembly. Proc Natl Acad Sci USA 2004; 101: 8664–8669.
Li JB, Gerdes JM, Haycraft CJ, et al. Comparative genomics identifies a flagellar and basal body proteome that includes the BBS5 human disease gene. Cell 2004; 117: 541–552.
Badano JL, Ansley SJ, Leitch CC, Lewis RA, Lupski JR, Katsanis N . Identification of a novel Bardet-Biedl syndrome protein, BBS7, that shares structural features with BBS1 and BBS2. Am J Hum Genet 2003; 72: 650–658.
Nishimura DY, Swiderski RE, Searby CC, et al. Comparative genomics and gene expression analysis identifies BBS9, a new Bardet-Biedl syndrome gene. Am J Hum Genet 2005; 77: 1021–1033.
Chiang AP, Beck JS, Yen HJ, et al. Homozygosity mapping with SNP arrays identifies TRIM32, an E3 ubiquitin ligase, as a Bardet-Biedl syndrome gene (BBS11). Proc Natl Acad Sci USA 2006; 103: 6287–6292.
Kyttälä M, Tallila J, Salonen R, et al. MKS1, encoding a component of the flagellar apparatus basal body proteome, is mutated in Meckel syndrome. Nat Genet 2006; 38: 155–157.
Arts HH, Doherty D, van Beersum SE, et al. Mutations in the gene encoding the basal body protein RPGRIP1L, a nephrocystin-4 interactor, cause Joubert syndrome. Nat Genet 2007; 39: 882–888.
Blacque OE, Reardon MJ, Li C, et al. Loss of C. elegans BBS-7 and BBS-8 protein function results in cilia defects and compromised intraflagellar transport. Genes Dev 2004; 18: 1630–1642.
Hildebrandt F, Otto E, Rensing C, et al. A novel gene encoding an SH3 domain protein is mutated in nephronophthisis type 1. Nat Genet 1997; 17: 149–153.
Olbrich H, Fliegauf M, Hoefele J, et al. Mutations in a novel gene, NPHP3, cause adolescent nephronophthisis, tapeto-retinal degeneration and hepatic fibrosis. Nat Genet 2003; 34: 455–459.
Mollet G, Salomon R, Gribouval O, et al. The gene mutated in juvenile nephronophthisis type 4 encodes a novel protein that interacts with nephrocystin. Nat Genet 2002; 32: 300–305.
Otto EA, Loeys B, Khanna H, et al. Nephrocystin-5, a ciliary IQ domain protein, is mutated in Senior-Loken syndrome and interacts with RPGR and calmodulin. Nat Genet 2005; 37: 282–288.
Valente EM, Silhavy JL, Brancati F, et al. Mutations in CEP290, which encodes a centrosomal protein, cause pleiotropic forms of Joubert syndrome. Nat Genet 2006; 38: 623–625.
Otto EA, Trapp ML, Schultheiss UT, Helou J, Quarmby LM, Hildebrandt F . NEK8 mutations affect ciliary and centrosomal localization and may cause nephronophthisis. J Am Soc Nephrol 2008; 19: 587–592.
Ruiz-Perez VL, Ide SE, Strom TM, et al. Mutations in a new gene in Ellis-van Creveld syndrome and Weyers acrodental dysostosis. Nat Genet 2000; 24: 283–286.
Galdzicka M, Patnala S, Hirshman MG, et al. A new gene, EVC2, is mutated in Ellis-van Creveld syndrome. Mol Genet Metab 2002; 77: 291–295.
Tallila J, Jakkula E, Peltonen L, Salonen R, Kestila M . Identification of CC2D2A as a Meckel syndrome gene adds an important piece to the ciliopathy puzzle. Am J Hum Genet 2008; 82: 1361–1367.
Noor A, Windpassinger C, Patel M, et al. CC2D2A, encoding a coiled-coil and C2 domain protein, causes autosomal-recessive mental retardation with retinitis pigmentosa. Am J Hum Genet 2008; 82: 1011–1018.
Eley L, Gabrielides C, Adams M, Johnson CA, Hildebrandt F, Sayer JA . Jouberin localizes to collecting ducts and interacts with nephrocystin-1. Kidney Int 2008; 74: 1139–1149.
Ou G, Blacque OE, Snow JJ, Leroux MR, Scholey JM . Functional coordination of intraflagellar transport motors. Nature 2005; 436: 583–587.
Kim JC, Badano JL, Sibold S, et al. The Bardet-Biedl protein BBS4 targets cargo to the pericentriolar region and is required for microtubule anchoring and cell cycle progression. Nat Genet 2004; 36: 462–470.
Kim J, Krishnaswami SR, Gleeson JG . CEP290 interacts with the centriolar satellite component PCM-1 and is required for Rab8 localization to the primary cilium. Hum Mol Genet 2008; 17: 3796–3805.
Kulaga HM, Leitch CC, Eichers ER, et al. Loss of BBS proteins causes anosmia in humans and defects in olfactory cilia structure and function in the mouse. Nat Genet 2004; 36: 994–998.
Tan PL, Barr T, Inglis PN, et al. Loss of Bardet Biedl syndrome proteins causes defects in peripheral sensory innervation and function. Proc Natl Acad Sci USA 2007; 104: 17524–17529.
Davis RE, Swiderski RE, Rahmouni K, et al. A knockin mouse model of the Bardet-Biedl syndrome 1 M390R mutation has cilia defects, ventriculomegaly, retinopathy, and obesity. Proc Natl Acad Sci USA 2007; 104: 19422–19427.
Nishimura DY, Fath M, Mullins RF, et al. Bbs2-null mice have neurosensory deficits, a defect in social dominance, and retinopathy associated with mislocalization of rhodopsin. Proc Natl Acad Sci USA 2004; 101: 16588–16593.
Eichers ER, Abd-El-Barr MM, Paylor R, et al. Phenotypic characterization of Bbs4 null mice reveals age-dependent penetrance and variable expressivity. Hum Genet 2006; 120: 211–226.
Weatherbee SOP, Alcorn H, Niswander L, Anderson K . Kerouac is required for proper cilia formation and patterning of multiple tissues. Dev Biol 2008; Program/Abstract no. 33.
Lowe LA, Supp DM, Sampath K, et al. Conserved left-right asymmetry of nodal expression and alterations in murine situs inversus. Nature 1996; 381: 158–161.
Mochizuki T, Saijoh Y, Tsuchiya K, et al. Cloning of inv, a gene that controls left/right asymmetry and kidney development. Nature 1998; 395: 177–181.
Zerres K, Rudnik-Schöneborn S, Deget F . Childhood onset autosomal dominant polycystic kidney disease in sibs: clinical picture and recurrence risk. German Working Group on Paediatric Nephrology (Arbeitsgemeinschaft für Pädiatrische Nephrologie). J Med Genet 1993; 30: 583–588.
Germino GG, Somlo S . Inherited diseases of the kidney. Curr Opin Nephrol Hypertens 1993; 2: 430–440.
Tobin JL, Beales PL . Restoration of renal function in zebrafish models of ciliopathies. Pediatr Nephrol 2008; 23: 2095–2099.
Bukanov NO, Smith LA, Klinger KW, Ledbetter SR, Ibraghimov-Beskrovnaya O . Long-lasting arrest of murine polycystic kidney disease with CDK inhibitor roscovitine. Nature 2006; 444: 949–952.
Eggenschwiler JT, Bulgakov OV, Qin J, et al. Mouse Rab23 regulates hedgehog signaling from smoothened to Gli proteins. Dev Biol 2006; 290: 1–12.
Author information
Authors and Affiliations
Corresponding author
Additional information
Disclosure: The authors declare no conflict of interest.
Rights and permissions
About this article
Cite this article
Tobin, J., Beales, P. The nonmotile ciliopathies. Genet Med 11, 386–402 (2009). https://doi.org/10.1097/GIM.0b013e3181a02882
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1097/GIM.0b013e3181a02882
Keywords
This article is cited by
-
1H, 13C, and 15N resonance assignments and solution structure of the N-terminal divergent calponin homology (NN-CH) domain of human intraflagellar transport protein 54
Biomolecular NMR Assignments (2024)
-
A local interplay between diffusion and intraflagellar transport distributes TRPV-channel OCR-2 along C. elegans chemosensory cilia
Communications Biology (2022)
-
Polymicrogyria, aventriculy, polydactyly, encephalocele, callosal agenesis (PAPEC): a new syndrome?
Child's Nervous System (2022)
-
Hypothalamic primary cilium: A hub for metabolic homeostasis
Experimental & Molecular Medicine (2021)
-
A novel splice site mutation in the SDCCAG8 gene in an Iranian family with Bardet–Biedl syndrome
International Ophthalmology (2021)
